Sophie Power — Year 3
Mentor: Garima Wilson
Introduction
Lichens are complex, mutualistic symbiotic organisms that function as integrated biological systems rather than single-species entities. Traditionally defined as an association between a fungal partner and a photosynthetic organism, lichens are now understood to be multi-partner ecosystems that include diverse microbial communities (Hawksworth & Grube, 2020). The dominant fungal partner, known as the mycobiont, forms the structural framework of the lichen and regulates water retention, mineral uptake, and protection from environmental stressors. The photosynthetic partner, or photobiont, is typically a green algae or a cyanobacterium and provides carbohydrates through photosynthesis (Duran-Nebreda & Valverde, 2023; Goodenough & Roth, 2021). Recent research has further demonstrated that lichens host additional microbial components, including basidiomycetous yeasts and bacterial assemblages, which contribute to nutrient cycling, stress tolerance, and overall symbiotic stability (Nguyen et al., 2023). This reconceptualization has led to the modern view of lichens as highly structured, self-regulating ecosystems rather than simple dual symbioses (Scharnagl et al., 2023). Lichens are also speculated to cover 8% of all surfaces on Earth (Drotos et al., 2024) and have over 14,000 known species (Lutzoni & Miadlikowska, 2009), making them one of the most abundant types of organisms.
The internal structure of a lichen thallus is highly organized and reflects its ecological function. Most lichens possess a layered anatomy consisting of an upper cortex, a photobiont layer, a medulla, and, in many species, a lower cortex (Goodenough & Roth, 2021). The upper cortex is composed of densely packed fungal hyphae and serves as a protective barrier against ultraviolet radiation, desiccation, and mechanical damage. Beneath this layer lies the photobiont zone, where algal or cyanobacterial cells are embedded among fungal hyphae to optimize light exposure while maintaining moisture availability. The medulla, composed of loosely interwoven hyphae, facilitates gas exchange and water storage. Because lichens lack true roots, vascular tissue, and a protective cuticle, they rely almost entirely on atmospheric deposition for water and nutrients, making them particularly sensitive to airborne contaminants (Thakur et al., 2024). This study aims to observe lichens within the Greater Vancouver area and investigate any correlation between the air quality and overall lichen health.
Lichens are commonly classified based on external morphology into three primary growth forms: crustose, foliose, and fruticose. Crustose lichens form thin, tightly adhered crusts that cannot be removed from their substrate without damage, while foliose lichens exhibit flattened, leaf-like lobes with distinct upper and lower surfaces. Fruticose lichens display three-dimensional, branching or filamentous growth forms. In foliose and fruticose lichens, the lower cortex may produce anchoring structures known as rhizines, which secure the lichen to its substrate without penetrating it. These morphological distinctions are ecologically significant, as they influence surface area exposure, moisture retention, and physiological sensitivity to environmental stressors, including air pollution (Thakur et al., 2024). Fruticose and foliose lichens are generally more sensitive to atmospheric pollutants than crustose species due to their increased surface area and reduced protective tissue density.
Reproduction in lichens occurs through both sexual and asexual mechanisms, primarily mediated by the fungal partner. Sexual reproduction involves the formation of fruiting bodies, most commonly apothecia, which appear as disc- or cup-shaped structures on the surface of the thallus and produce fungal spores known as ascospores (Bowler & Rundel, 1975). Less commonly, flask-shaped reproductive structures called perithecia may be embedded within the thallus. While sexual reproduction promotes genetic diversity, successful establishment of a new lichen requires the fungal spore to encounter a compatible photobiont after dispersal, making this process relatively inefficient. Asexual reproduction allows for simultaneous dispersal of both symbiotic partners through specialized structures such as soredia and isidia. Soredia are powdery granules composed of algal cells enveloped by fungal hyphae, whereas isidia are small, finger-like outgrowths that detach from the thallus. These structures enhance colonization efficiency and play a significant role in local persistence and expansion of lichen populations (Bowler & Rundel, 1975; Kantelinen et al., 2022).
Bioindicators are organisms used to assess the health of an ecosystem, as their physiological condition reflects cumulative environmental exposure rather than short-term fluctuations (Thakur et al., 2024). Lichens are particularly valuable bioindicators because they integrate environmental stress over long periods, making them sensitive and reliable indicators of long-term change (Thakur et al., 2024). This utility stems in part from their characteristically slow life cycle, with growth rates often measured in millimeters per year and lifespans extending from decades to centuries (Sanders, 2014). Establishment requires a suitable substrate, sufficient moisture, appropriate light conditions, and the successful formation or maintenance of the symbiotic relationship. Once established, lichens gradually expand their surface area and produce reproductive structures when environmental conditions are favorable.
Numerous studies have demonstrated strong correlations between lichen diversity, abundance, and physiological condition and atmospheric pollution levels. Thakur et al. (2024) documented predictable declines in lichen diversity and structural complexity in polluted environments, while Yang et al. (2023) demonstrated the accumulation of heavy metals within lichen tissues in urban ecosystems, linking tissue damage and reduced reproductive capacity to elevated contamination. These findings support the use of lichens as effective biological monitors of air quality across diverse ecosystems.
Air pollution remains a major global public health concern, particularly in urban environments where vehicle emissions, industrial activity, and urban expansion contribute to elevated concentrations of harmful pollutants. The World Health Organization (2019) estimates that approximately 99% of the global population breathes air that exceeds recommended pollution thresholds, increasing the risk of respiratory and cardiovascular disease, impaired development in children, and reduced life expectancy. The Greater Vancouver area presents a compelling context for this concern. As one of Canada’s most populous and rapidly growing metropolitan regions (Huang et al., 2025), Greater Vancouver has experienced significant urban expansion driven by population growth, intensified vehicular traffic, and industrial activity across sectors including port operations, construction, and natural gas processing. These pressures produce spatially variable air quality conditions, with pollution levels differing substantially between densely urbanized cores, residential neighbourhoods, and the natural areas, such as Pacific Spirit Park and Stanley Park, which border the city (Huang et al., 2025). Despite the scale of these challenges, conventional air quality monitoring infrastructure remains limited in spatial resolution and is costly to build and maintain. This study addresses this gap by evaluating lichens as accessible, low-cost bioindicators of air quality across the Greater Vancouver area, examining whether observable differences in lichen structure, reproduction, and overall health correspond to expected pollution gradients between natural, residential, and urban sites.
Methodology
Study Design
This study employed a field-based observational survey to evaluate lichen health and diversity as indicators of air quality across the Greater Vancouver area. Rather than measuring atmospheric pollutants directly, this approach assesses biologically-integrated responses to long-term air exposure through measurable lichen traits. Observations were conducted at multiple sites representing a gradient of environmental conditions, from relatively undisturbed natural areas to highly urbanized locations with increased traffic density.
Site Selection
Study sites were selected to capture variation in potential air pollution exposure. Natural environments such as Pacific Spirit Park and Stanley Park were chosen as lower-impact reference sites due to reduced traffic and higher vegetation density. Residential neighborhoods represented moderate exposure levels, while urban locations such as the downtown Vancouver Public Library branch represented areas with high human activity and vehicular emissions. All sites were surveyed using consistent observation protocols to ensure comparability.
Sampling Methods
Within each site, multiple lichen samples were identified on stable substrates such as tree bark or rock surfaces. Sampling three individuals per site ensured data continuity if individual specimens became damaged, detached, or otherwise unsuitable for observation. Lichens were not removed from their substrates and all observations were conducted on site to minimize disturbance.
Method of Observation and Measured Traits
For each lichen sample, a standardized set of traits were observed and recorded, as these characteristics are known to reflect physiological stress and long-term exposure to airborne pollutants. In the field, the data was documented in a physical log book, and when back from the field, data was catalogued digitally.
Necrosis was assessed by identifying areas of dead or dying tissue, characterized by darkened, brittle, or fragmented regions of the thallus. The presence and extent of the necrosis were used as indicators of physiological stress, as increased pollutant exposure disrupts cellular processes and can lead to tissue death. Necrosis was measured as a percentage of its area coverage.
Area coverage was measured in square millimeters (mm²) by using a measuring grid to see the surface area occupied by each lichen thallus. Reduced surface coverage or shrinking thalli were interpreted as signs of unfavourable environmental conditions, while stable or expanding coverage suggested healthier growth and better air quality.
Species number was recorded at each site by counting the number of distinct lichen species present within the sampling area. Higher species richness was interpreted as indicative of cleaner air, as many lichen species, particularly foliose and fruticose forms, are sensitive to atmospheric pollutants. Sites dominated by few, pollution-tolerant species were interpreted as experiencing poorer air quality.
The photobiont type was inferred based on thallus coloration and texture. Darker, often bluish or brownish lichens were identified as likely containing cyanobacteria while brighter green lichens were inferred to contain green algal photobionts. Cyanobacterial lichens are generally more sensitive to pollution and environmental stressors, making their presence or absences a relevant indicator of air quality.
The type of reproduction was documented by observing reproductive structures present on the thallus. Sexual reproduction was identified by the presence of apothecia or perithecia, while asexual reproduction was identified through structures such as soredia or isidia. Reduced or absent reproductive structures were interpreted as signs of environmental stress, as lichens under poor conditions prioritize survival over reproduction.
The length and thickness of stalks and reproductive structures, where present, were measured in millimeters (mm) with a ruler. Reduced size or malformed structures were interpreted as signs of impaired growth, while well developed structures suggested favourable environmental conditions.
Data Recording and Documentation
All observations were recorded systematically in a data log book when on site, and in standardized data sheets when no longer on site. Photographic documentation was collected for each sample to verify observations, support species identification, and allow for later comparisons. Measurements were taken using measuring grids to minimize observer error.
Results
Lichen Surface Area
Surface area coverage varied substantially across the three study sites and showed divergent trajectories over the observation period. At the park site, the mean surface area across three lichen samples increased from an average of 51.61 cm² on the first observation to 64.52 cm² by the last observation (Figure 1), representing a 25.0% increase over the observation period. This upward trajectory was consistent across all three samples at this site, with Sample 1 (fruticose morphology) increasing from 38.17 to 51.61 cm², Sample 2 (foliose morphology) from 70.97 to 83.87 cm², and Sample 3 (foliose morphology) from 45.16 to 58.06 cm².
At the residential neighbourhood site, mean surface area increased from 17.23 cm² to 23.68 cm² over the same period, representing a 37.5% increase, a larger average change relative to samples from the park site, though the absolute dimensions remained considerably smaller. Growth was uneven across samples: Sample 1 (foliose morphology) grew from 6.45 to 19.35 cm², Sample 2 (crustose morphology) showed modest growth from 38.71 to 45.16 cm², and Sample 4 (foliose morphology) remained static at 6.45 cm² throughout the study period. At the city site, mean surface area remained entirely static at 8.58 cm² across all observation dates (Figure 6). All three samples maintained their initial measurements without any measurable growth.
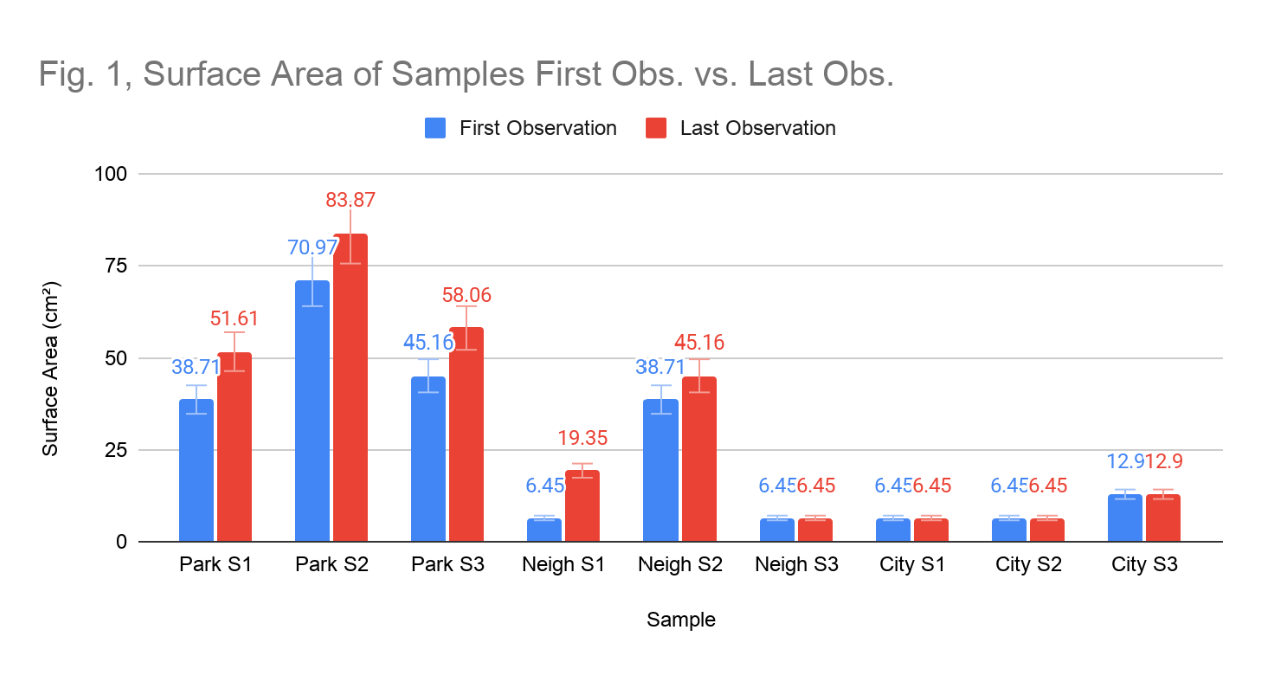
Figure 1: Surface Area of Samples First Obs. vs. Last Obs. Comparison of the surface area (cm²) of each lichen sample on the first observation date (blue) and on the last observation date (red).
Necrosis
Necrosis levels differed among sites, and at the city location, increased substantially over the observation period. At the park site, necrosis remained stable and relatively low throughout the study. Samples 1 and 2 maintained consistent necrosis at 10% and 5% respectively across all observation dates, while Sample 3 exhibited no evidence of necrosis at any point (Figure 2). The mean site-level necrosis was 5.0%, with no change observed. At the neighbourhood site, necrosis was similarly moderate but showed evidence of progression in certain samples. Samples 1 and 3 initially showed no necrosis (Figure 2), but by the final observation, Sample 1 had increased by 5% and Sample 3 remained at a persistent 15%. Sample 2 of the neighbourhood site, showed no necrosis for the first three observations, but towards the end of the study had necrotized by 5%. This mean necrosis across the neighbourhood site was 5.42% (Figure 3).
Necrosis at the city site was the highest recorded across all sites and showed clear progression. Sample 1 began with 10% necrosis and increased to 25% by the end of the study. Sample 3 similarly progressed from 10% to 15% necrosis over the study period (Figure 4). Only Sample 2 showed no necrosis at an observation date. The mean necrosis at the city site was 10.83%, more than double the park value, and the pattern of increase over time suggests ongoing, active physiological deterioration rather than pre-existing stable damage.
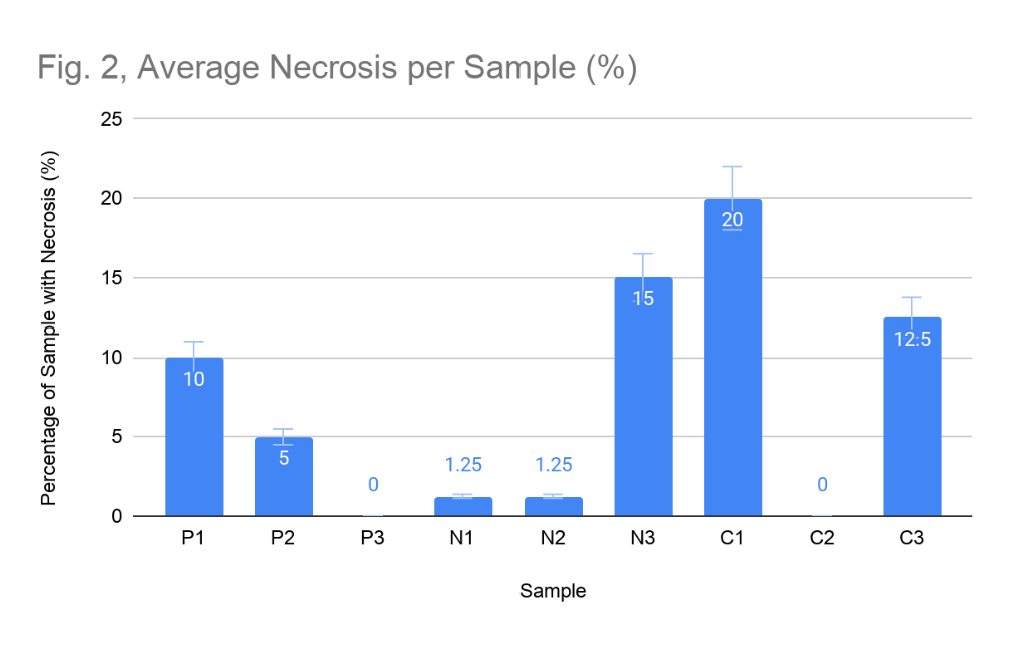
Figure 2: Average Necrosis per Sample (%). Average percentage of necrosis in each lichen sample (P = park, N = neighbourhood, C = city), measured as the percentage of the sample that was necrotic or damaged.
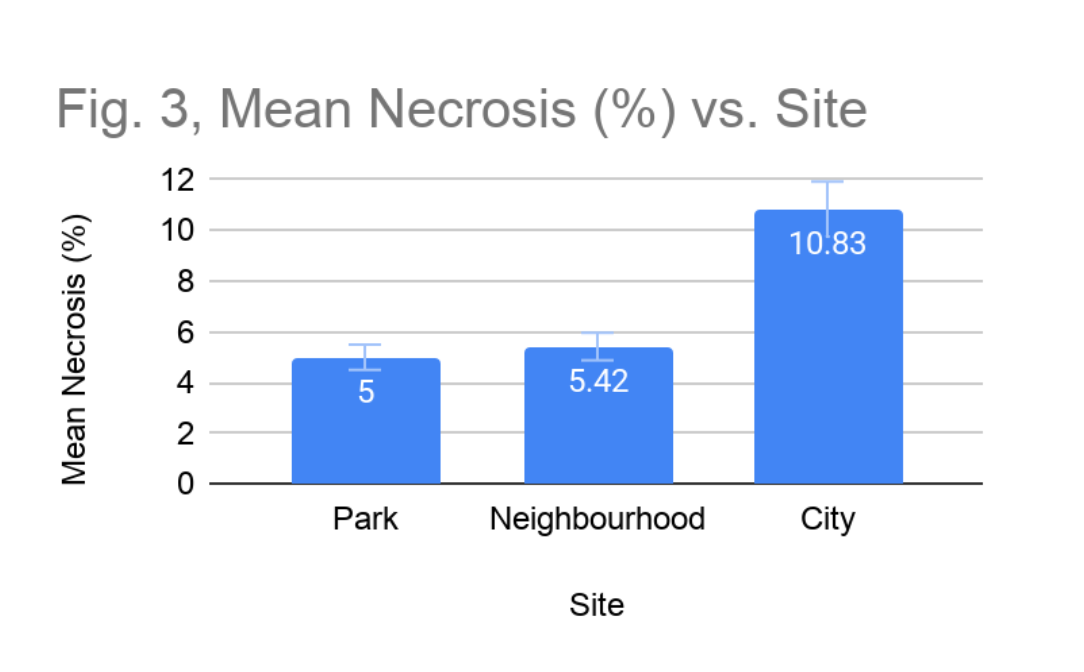
Figure 3: Mean Necrosis (%) vs. Site. Mean necrosis of each sample site, calculated by averaging the final percentage of necrosis across all lichens at each site.
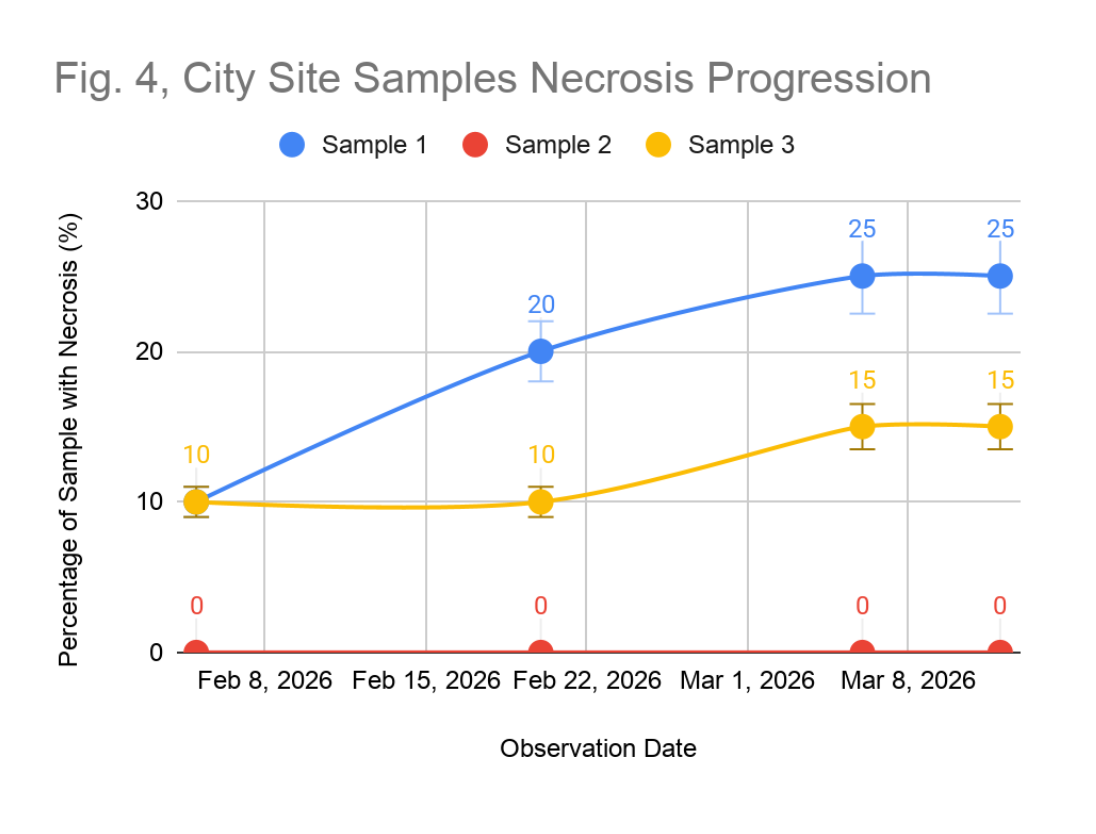
Figure 4: City Site Samples Necrosis Progression. Progression of necrosis (%) across city site samples (Sample 1, blue; Sample 2, red; Sample 3, yellow) from February 8 to March 8, 2026.
Species Composition and Reproductive Mode
Species composition and reproductive strategies differ distinctly across sites, following a pattern consistent with established pollution-sensitivity gradients. At the park site, the samples community included both foliose and fruticose growth forms, with all three samples exhibiting sexual reproduction and a cyanobacterial photobiont. No crustose lichens were sampled at this site. At the neighbourhood site, sampled species were foliose and crustose (Figure 5). Notably, the foliose samples at this site reproduced asexually, while the crustose sample reproduced sexually. Photobiont composition was mixed, with the foliose samples hosting cyanobacteria and the crustose sample hosting green algae. At the city site, the samples community was crustose and foliose, with all three samples reproducing asexually and hosting cyanobacteria photobionts. Stalk thickness was uniformly zero at both park and city sites, while one sample in the neighbourhood site showed a stalk thickness of 1 mm.
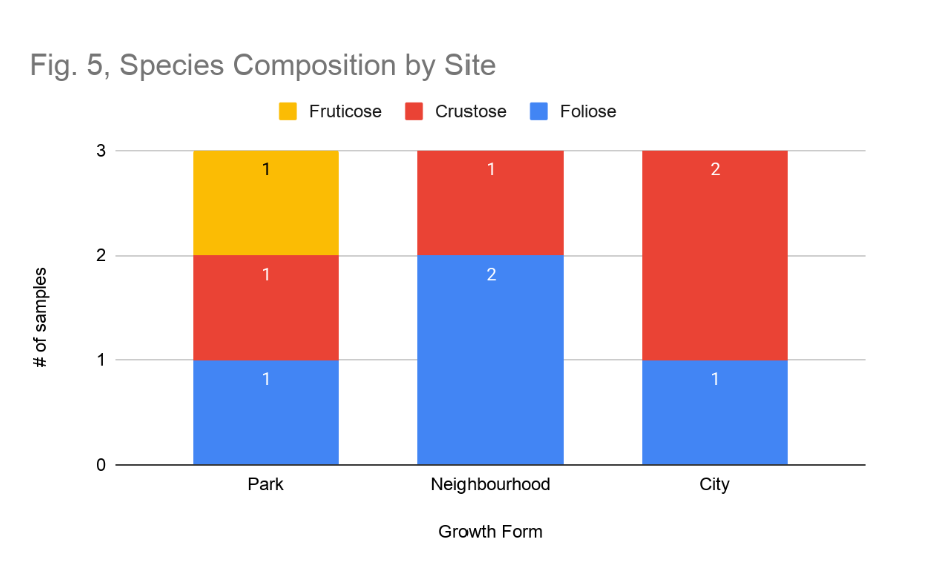
Figure 5: Species Composition by Site. Number of fruticose (yellow), crustose (red), and foliose (blue) lichen samples at each site.
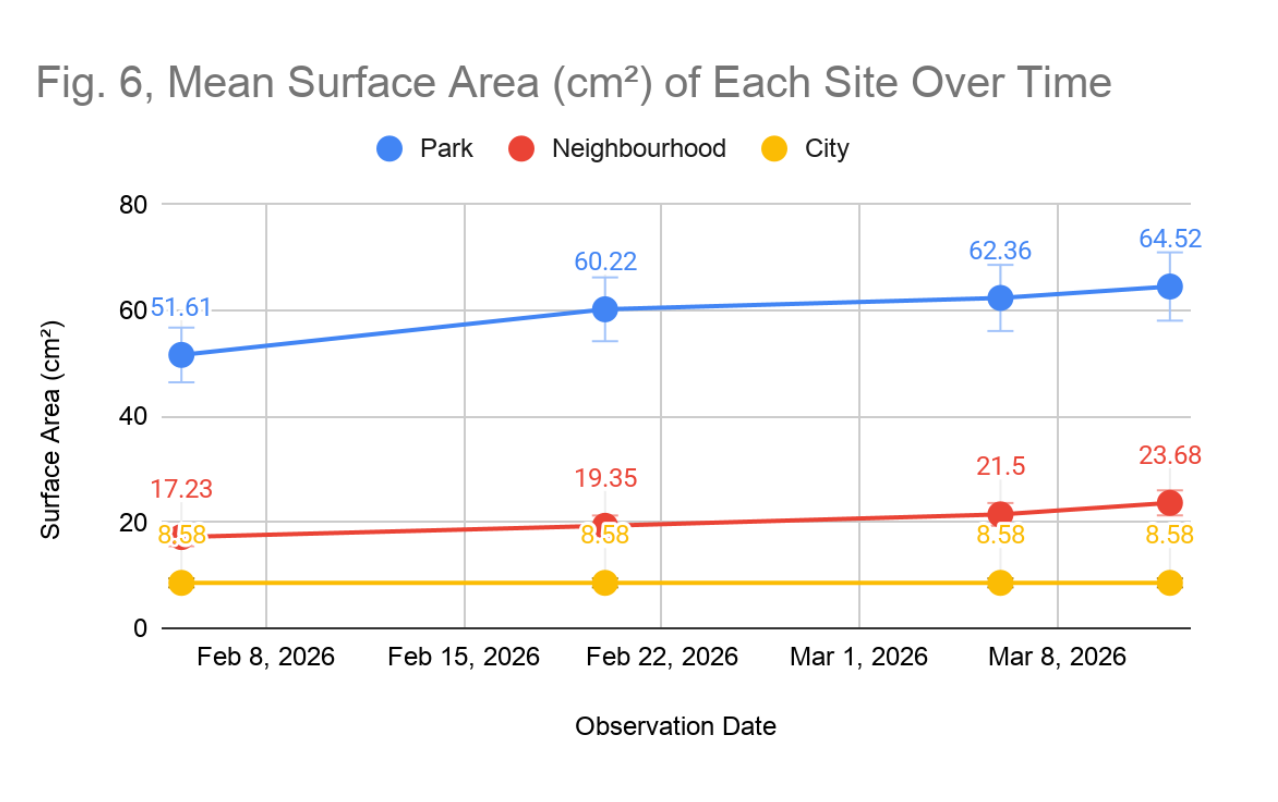
Figure 6: Mean Surface Area (cm²) of Each Site Over Time. Mean surface area of lichen samples at the park (blue), neighbourhood (red), and city (yellow) sites across five observation dates from February 8 to March 8, 2026.
Discussion
Surface Area as an Indicator of Lichen Vitality
The surface area data reveal a clear urban gradient in lichen growth capacity, with the park specimens exhibiting active and consistent thallus expansion, neighbourhood specimens showing limited and uneven growth, and city specimens exhibiting complete growth cessation over the observation period. These findings are consistent with the well-established relationship with urbanization and lichen vitality described in literature. Thakur et al. (2024) identified reductions in lichen surface coverage as a reliable indicator of pollutant-induced physiological impairment, as atmospheric contaminants, particular nitrogen oxides, sulfur oxide, and particulate matter from vehicular emissions. They disrupt photosynthetic efficiency in photobionts, thereby reducing the carbohydrate supply available for thallus growth. Because lichens acquired nutrients entirely through atmospheric deposition and lack vascular tissue to buffer environmental inputs, even moderate elevations in pollutant concentrations can suppress growth rates measurably (Thakur et al., 2024).
The complete absence of growth at the city site is particularly notable. While lichen growth rates are characteristically slow, the park specimens showed measurable expansion within the observation period of six weeks. This suggests the site conditions were sufficiently favourable to permit net positive thallus growth. The inability of the city specimens to achieve any measurable growth over the same period of time implies that environmental conditions at the city site are either insufficient for net carbon fixation or that metabolic resources are being diverted toward stress response pathways at the expense of growth. This interpretation is supported by the concurrent necrosis data from the same site, which indicate active cellular damage. It should be noted, however, that baseline differences in the surface area between sites, with the park lichens averaging 56.98 cm² compared to 8.58 cm² at the city site (Figure 6), may partly reflect pre-existing colonization histories and substrate availability rather than solely pollution effects, and this confounding factor should be considered when interpreting the comparative data.
Necrosis as a Marker of Physiological Stress
The necrosis data provides the most compelling evidence of an air quality gradient across the study sites. The inverse relationship between environmental urbanization and lichen tissue health, with mean necrosis values of 5.0%, 5.42%, and 10.83% at the park, neighbourhood, and city sites respectively (Figure 3), is consistent with mechanisms of pollutant-induced cellular damage documented in the lichen bioindicator literature. Yang et al. (2023) demonstrated that heavy metal accumulation within lichen thalli, a consequence of their passive uptake of atmospheric deposition, leads to oxidative stress, disruption of enzymatic activity, and ultimately cell death, manifesting as the visible tissue darkening and fragmentation characteristic of necrosis. Elevated concentrations of nitrogen-based pollutants, common in urban environments with high traffic density, can further acidify the lichen thallus surface and denature the cellular machinery of the photobiont, accelerating tissue breakdown (Thakur et al., 2024).
Of particular significance is the progression of necrosis at the city site (Figure 4). Sample 1 progressed from 10% to 25% necrosis during the study, representing a 150% increase in affected tissue area. This rate of deterioration is not consistent with pre-existing chronic damage, which would be expected to remain relatively stable over short observation intervals, but rather suggests acute and ongoing physiological stress. This finding corroborates the hypothesis that the city site represented a high-pollutant exposure environment in which lichen health is acutely and continuously compromised. By contrast, the stable and low necrosis levels observed at the park site, despite the presence of necrosis in two of three samples suggest that background-level tissue damage exists even in relatively clean environments, possibly attributable to natural senescence, mechanical damage, or localised moisture variability rather than air quality alone. This distinction between stable background necrosis and actively progressing tissue death is an important interpretive nuance that should be considered in biomonitoring applications.
Species Composition, Growth Form, and Pollution Tolerance
The compositional shift in lichen growth forms across the pollution gradient provides additional support for the air quality hypothesis (Figure 5). The presence of fruticose lichens exclusively at the park site aligns with their well-documented status as the most pollution-sensitive growth form. This trait is attributed to their three-dimensional morphology, large surface area to volume ratio, and thin cortical tissue offering limited protection against pollutant deposition (Thakur et al., 2024). The dominance of crustose morphotypes at the neighbourhood and city sites mirrors patterns observed across multiple urban biomonitoring studies, in which pollution-tolerant crustose species persist in conditions that exclude more sensitive foliose and fruticose taxa. The tightly appressed thallus of crustose lichens minimizes atmospheric exposure and limits the uptake of harmful compounds, conferring a relative advantage in degraded air quality environments.
The shift from sexual to asexual reproduction across the pollution gradient is likewise consistent with predictions derived from lichen stress physiology. Sexual reproduction in lichens requires the metabolic investment of producing apothecia or perithecia, a process dependent on sustained photosynthetic output and physiological stability (Bowler & Rundel, 1975). Under conditions of environmental stress, lichens are theorized to downregulate energetically costly reproductive processes in favour of maintenance and survival. All sampled lichens at the city site showed evidence of asexual reproduction and none exhibited apothecial development, while all park specimens reproduced sexually. These patterns suggest that city site lichens are operating under sufficient stress to preclude sexual reproductive investment. Kantelinen et al. (2022) notes that reproductive mode is tightly coupled to substrate stability and environmental quality, further supporting the interpretation that the shift toward asexual reproduction at urban sites reflects adverse conditions rather than species-specific reproductive strategy alone. It is worth noting, however, that the observed photobiont composition with cyanobacteria occurring at the neighbourhood site rather than being excluded from intermediate pollution environments does not fully conform to the expectation that cyanobacterial lichens should be restricted to cleaner conditions. This may reflect the relatively short observation window, possible misidentification of photobiont type based on coloration, or genuine site-specific tolerances that warrant further investigation.
Limitations and Methodological Considerations:
Several limitations constrain the strength of the conclusions that can be drawn from this study. The sample size of three lichens per site is small, limiting the statistical power to distinguish site level trends from individual variation. The six week observation window, while sufficient to capture short term changes in necrosis and growth, does not reflect the multi-year timescales over which lichen communities typically respond to and recover from pollution exposure (Sanders, 2014). Consequently, current species assemblages at each site may reflect historical air quality conditions rather than the present day pollution gradient, and year to year variability in weather patterns, including precipitation, temperature, and humidity, could independently influence the growth and necrosis metrics measured. Late winter and early spring conditions in Vancouver may impose seasonal constraints on lichen physiology that are unrelated to pollution, as moisture availability, light intensity, and temperature all influence lichen metabolic activity and could partly explain the growth patterns observed (Thakur et al., 2024).
The inference of photobiont type from thallus colouration, while methodologically practical for field surveys, introduces the possibility of misclassification, as colouration can vary with hydration state, substrate chemistry, and the presence of secondary lichen compounds unrelated to photobiont identity. Similarly, necrosis percentage was estimated visually from in person observations and image analysis rather than histological examination, which may introduce inter-observer variability. Future studies would benefit from direct atmospheric pollutant concentrations, to enable correlation analyses between specific pollutant loads and lichen health metrics, providing a more mechanistic link between air quality and the biological responses documented. Expanding the number of sites, sample replicates, and observation intervals would also substantially strengthen the conclusions that can be drawn regarding the utility of lichen as bioindicators in the Greater Vancouver urban environment.
Broader Implications
Despite these limitations, the results of this study broadly support the use of lichen assemblages as accessible, low-cost indicators or relative air quality in urban environments. The convergence of multiple independent lines of evidence like surface area growth rates, necrosis levels, growth form composition, and reproductive strategy, point in the same direction across the urbanization gradient lends confidence to the overall interpretation. The methodology employed here requires no specialised equipment beyond a measuring grid and standardised field protocol, making it replicable at a scale and potentially applicable in community science or educational contexts. Given the limited spatial resolution of conventional air quality monitoring infrastructure and the WHO’s (2019) estimate that the vast majority of the global population is exposed to air exceeding recommended pollution thresholds, bioindicator-based approaches such as this study offer a meaningful complement to instrumental monitoring. Further research integrating lichen biomonitoring with geospatial analysis of traffic density, industrial emissions, and prevailing wind patterns of the Greater Vancouver region could yield a fine-grained, biologically validated map of air quality variation across the city, with direct relevance to public health planning and urban environmental policy.
Appendix
| Obs. Date | S1 | S2 | S3 | S1 | S2 | S3 | S1 | S2 | S3 |
| Feb 5, 2026 | 38.71 | 70.97 | 45.16 | 6.45 | 38.71 | 6.45 | 6.45 | 6.45 | 12.9 |
| Feb 20, 2026 | 45.16 | 77.42 | 58.06 | 6.45 | 45.16 | 6.45 | 6.45 | 6.45 | 12.9 |
| Mar 6, 2026 | 51.61 | 77.42 | 58.06 | 12.9 | 45.16 | 6.45 | 6.45 | 6.45 | 12.9 |
| Mar 12, 2026 | 51.61 | 83.87 | 58.06 | 19.35 | 45.16 | 6.45 | 6.45 | 6.45 | 12.9 |
World Health Organisation. (2019, July 30). Air pollution. Who.int; World Health Organization: WHO. https://www.who.int/health-topics/air-pollution
Thakur, M., Bhardwaj, S., Kumar, V., & Rodrigo-Comino, J. (2024). Lichens as effective bioindicators for monitoring environmental changes: A comprehensive review. Total Environment Advances, 9(Volume 9), 200085. https://doi.org/10.1016/j.teadva.2023.200085
Yang, J., Oh, S.-O., & Hur, J. (2023). Lichen as Bioindicators: Assessing their Response to Heavy Metal Pollution in Their Native Ecosystem. Mycobiology, 51(5), 343–353. https://doi.org/10.1080/12298093.2023.2265144
Hawksworth, David L., and Martin Grube. “Lichens Redefined as Complex Ecosystems.” New Phytologist, vol. 227, no. 5, 2 June 2020, pp. 1281–1283, https://doi.org/10.1111/nph .16630.
Duran-Nebreda, Salva, and Sergi Valverde. “Composition, Structure and Robustness of Lichen Guilds.” Scientific Reports, vol. 13, no. 1, 25 Feb. 2023, p. 3295, www.nature.com/articles/s41598-023-30357-w , https://doi.org/10.1038/s41598-023-30357-w.
Nguyen, Ngoc-Hung, et al. “Biodiversity of Basidiomycetous Yeasts Associated with Cladonia Rei Lichen in Japan, with a Description of Microsporomyces Cladoniophilus Sp. Nov.” Journal of Fungi, vol. 9, no. 4, 14 Apr. 2023, pp. 473–473, https://doi.org/10.3390/jof9040473 . Accessed 10 May 2025.
Goodenough, Ursula, and Robyn Roth. “Lichen 2. Constituents.” Algal Research, vol. 58, 1 Oct. 2021, p. 102356, http://www.sciencedirect.com/science/article/abs/pii/S2211926421001752, https://doi.org/10.1016/j.algal.2021.102356. Accessed 24 Aug. 2021.
Kantelinen, Annina, et al. “Lichen Speciation Is Sparked by a Substrate Requirement Shift and Reproduction Mode Differentiation.” Scientific Reports, vol. 12, no. 1, 30 June 2022, p. 11048, pubmed.ncbi.nlm.nih.gov/35773369/ , https://doi.org/10.1038/s41598-022-14970-9
Scharnagl, Klara, et al. “The Coming Golden Age for Lichen Biology.” ScienceDirect, vol. 33, no. 11, 1 June 2023, pp. R512–R518, https://doi.org/10.1016/j.cub.2023.03.054 .
Bowler, Peter A, and Philip W Rundel. “Reproductive Strategies in Lichens.” Botanical Journal of the Linnean Society, vol. 70, no. 4, 1 June 1975, pp. 325–340, https://doi.org/10.1111/j.1095-8339.1975.tb01653. x. Accessed 24 Apr. 2023.
Ali, Farak, et al. “Commercial and Traditional Uses of Lichen.” Chemistry, Biology and Pharmacology of Lichen, 7 June 2024, pp. 125–136, https://doi.org/10.1002/9781394190706.ch10 . Accessed 31 Jan. 2026.
Sanders, William B. “Complete Life Cycle of the Lichen Fungus Calopadia Puiggarii (Pilocarpaceae, Ascomycetes) Documented in Situ: Propagule Dispersal, Establishment of Symbiosis, Thallus Development, and Formation of Sexual and Asexual Reproductive Structures.” American Journal of Botany, vol. 101, no. 11, Nov. 2014, pp. 1836–1848, https://doi.org/10.3732/ajb.1400272