Jayden Zhang — Year 2, Life Science
Abstract
Ultraviolet radiation is one of the most powerful and common mutagens on earth, capable of causing direct and indirect DNA damage through lesions and oxidative stress, which can ultimately lead to carcinogenesis. The objective of this study was to observe the susceptibility of E. coli K12 to UVB-induced damage across varying exposure durations, using a UVB reptile lamp designed to mimic sunlight. Bacterial cultures were spread-plated at a 10⁻⁴ dilution and immediately exposed for 0 (control), 30, 60, or 90 minutes, followed by immediate incubation, with colony counts used to measure effects. The results obtained show a strong correlation between a longer period of UV exposure and greater damage to the bacteria, observed through a lower percentage of control colonies remaining after longer periods of exposure: on average, after 30, 60, and 90 minutes, 64.7%, 39.0%, and 21.4% of colonies survived respectively, in proportion to each of their control groups.
Introduction
Ultraviolet (UV) light is a type of invisible electromagnetic radiation, emitted mainly from the sun, but also found in a plethora of manmade tools. It has a wavelength between 100 and 400 nanometers and is classified into 3 types based on energy of the waves, which is inversely related to the wavelength. UVA and UVB, are two types which we interact with on a daily basis, but there is also UVC, which is mostly absorbed and filtered out by our atmosphere. The rays of ultraviolet light that make it through the atmosphere are some of the most powerful carcinogenic agents that are capable of altering cell behavior by inducing mutagenic and cytotoxic lesions (Rastogi et al., 2010) and are the main cause of skin cancer cases around the world. UV light causes skin cancer by hitting our cells and breaking DNA, creating mutations and therefore raising possibilities of carcinogenesis (Kciuk et al., 2020). This radiation can also increase the chance of developing other cancers. Understanding UV radiation and its effect on life is therefore crucial for public health and biological research, and studying these effects can provide insight into the biological mechanisms of cancer and mutations to further research on cancer prevention.
The main biological harm of UV light is its ability to cause damage to cellular DNA, with UVB being the type of UV that causes the most harm to humans on a daily basis. UVB makes up 10% of the UV light we are exposed to, and its rays contain enough energy to directly damage cellular DNA in tissue near the surface of our skin. Furthermore, exposure of genomic DNA to UV light leads to the formation of many different types of damage (Kciuk et al., 2020), making the harm caused by UV exposure more difficult to precisely quantify. Two prominent types of direct DNA damage caused by UV exposure are cyclobutane pyrimidine dimers (CPDs) and 6-4 photoproducts. CPDs occur when ultraviolet light hits DNA and causes “adjacent pyrimidine bases [to become] covalently linked” (Kciuk et al., 2020), via the C5 and C6 carbon atoms on the DNA structure. 6-4 photoproducts occur when “a noncyclic bond is formed between the C6 and C4 atoms of pyrimidine residues” (Kciuk et al., 2020). These types of direct damage affect DNA’s double helix structure, which can block replication and various essential pathways in the cell, like immune pathways and replication pathways, leading to mutations and ultimately carcinogenesis.
Indirect damage to DNA from UV exposure occurs “mostly through the accumulation of 8-oxo-7,8-dihydro-2′-deoxyguanosine” (Zhivagui et al., 2023), also known as oxidative stress, and is mostly caused by UVA, which makes up 90% of the UV light we are exposed to. This type of damage does not directly impact the DNA, however due to the limited ability for human skin cells to deal with the multitudes of UV damage, UVA exposure is responsible for the accumulation of damage in skin cells weakening the cell’s ability to further repair damage (Leccia et al., 2019). If one cell turns cancerous, it does not mean you have cancer, it is the accumulation of these various damages on your skin via daily exposure and the build up of indirect and direct damages that truly causes skin cancer.
When DNA is damaged, cells will try to repair it using a plethora of methods, however cells can prioritize their own survival over the potential mutation and improper repair of the DNA, leading to mutations compounded by the original damage. Additionally, many animals and plants will have different systems of repair to account for the same types of damage. There are two main methods of repair for ultraviolet associated DNA damage in the human body: nucleotide excision and base excision repair. The nucleotide excision repair pathway, or NER, is specialized in repairing 6-4 photoproducts, but can perform poorly when attempting to repair CPDs (Leccia et al., 2019). The BER or base excision repair pathway is the predominant DNA repair pathway against base lesions caused by damaging byproducts called reactive oxygen species (Rastogi et al., 2010), also understood as oxidative stress. This pathway accounts for the indirect damage caused by UVA on DNA, but can also repair CPDs and photoproducts.
Humans are not the only organisms whose DNA is affected by UV light, however. Bacteria and plants have also evolved to have specific enzymes and pathways responsible for repairing the damage caused to their DNA by UV radiation, and are much more accessible for research than human cells. A prominent UV recovery mechanism in plants and bacteria is photorepair, which centers around using a well-conserved enzyme called photolyase. This enzyme uses the energy of visible blue-light to protect the genome from UV light induced harm (Rastogi et al., 2010), and can “greatly [enhance] the rate of removal of dimers, although, in the absence of photoreactivating (blue) light, dimers are slowly eliminated from bulk DNA and 6-4PPs are generally observed to be repaired more quickly than CPDs”, due to the enzymes not having energy. This pathway is currently being used in research for a photolyase containing “photorepair” skin product to revert the effects of DNA damage via UV over time, that should be used along with “photoprotection” products like sunscreen. Other methods that plants and bacteria use to negate UV damage include “shielding by flavonoids and phenolic compounds”, with these compounds simply acting as physical barriers, absorbing the UV light before it can damage the DNA. Further understanding how bacteria and plants resist UV light can be greatly beneficial not only for research, but also for the creation of products similar to the “photorepair” concept.
The primary cause of skin cancer around the world is UV exposure, more specifically UVB exposure, with a study showing UV exposure as the primary risk factor for melanoma, and histories of sunburn and frequent tanning correlating to higher cancer risk (Chang et al., 2022). This is because skin cancer develops from UV damage via accumulation of both indirect and direct damages, as unrepaired lesions and damage accumulation has different effects on your body depending on quantity and severity of exposure, with effects including breaking tumor suppressing pathways, and damaging proto-oncogenes. Understanding these effects is important to understanding the mutation and development of skin cancer in your body.
The importance of understanding the interaction of bacteria and UV radiation and the impact that exposure time and frequency has on mutations was discussed above as important places for research to be conducted, which is why I chose to do my research project on this specifically. The objective of this study is to understand the susceptibility of E. coli K12 to mutation from a single UV light exposure of different times using a UV reptile lamp designed to mimic the sun, with the objective of observing different colony traits than those of the control groups.
Ultraviolet radiation presents a significant environmental threat to humans because of its ability to cause skin cancer and disrupt people’s lives. Electromagnetic radiation can cause direct and indirect DNA damage through lesions or oxidative stress, and can ultimately cause carcinogenesis and skin cancer through the accumulation of damage. Humans, plants, and bacteria have many different mechanisms for the repair and negation of this damage, including excision pathways, enzyme repairs, and physical barriers. This aim of this study is to correlate UV exposure with damage to DNA and mutation of bacteria to further strengthen the understanding of DNA damage via UV radiation.
Materials and Methods
This study used basic bacterial plating materials for E-Coli K12, including petri dishes, nutrient agar, an incubator, and tools for spread-plating the bacteria. The bacteria were spread plated via a standard procedure.

Figure 1: Standard spread plating procedure (image from https://microbenotes.com/spread-plate-technique/)
The sunlight exposure was simulated using an opaque box made of polycarbonate plastic (Polymershapes) and a UVB reptile bulb (Exoterra UVB 100 light). The exposure groups were separated by length of exposure time, with the test groups being 30 minutes, 60 minutes, and 90 minutes, each beginning exposure immediately after plating and entering the incubator immediately after their exposure period. The control group was simply placed directly into the incubator after being plated. The plates were then left for one day, then put into the fridge until further analysis.

Figure 2: The experimental set up that was employed. The polycarbonate box combined with the aluminum sheet prevents UVB light from reaching other’s eyes, and these materials are fire resistant to avoid catching on fire from the UV lamp’s heat.
For observation, the plates were taken out of the fridge after their growth period, and the individual colonies of bacteria were counted. These results were then compared to each other and their respective exposure periods.
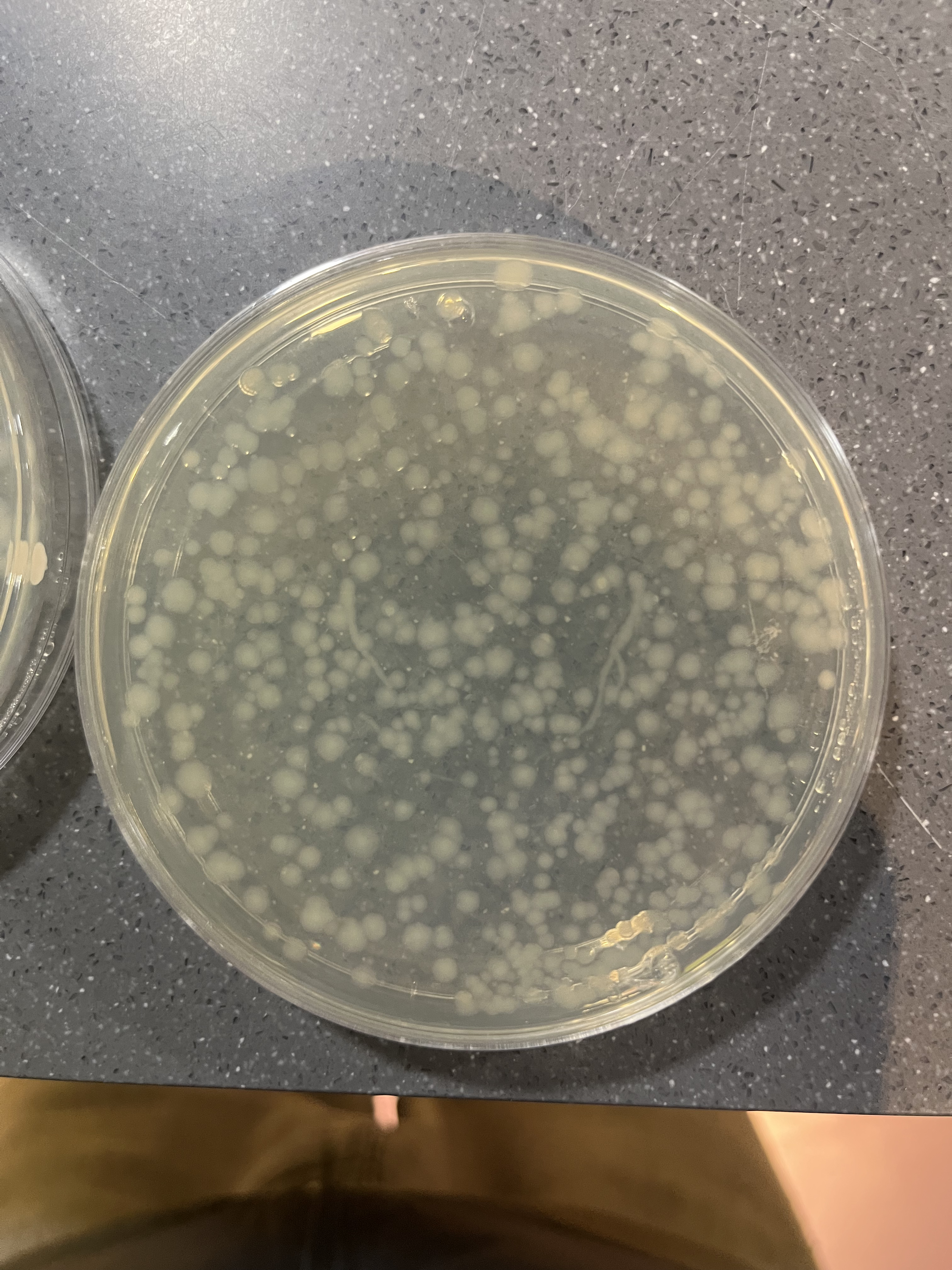
Figure 3: Inoculated E. coli plate post-incubation
To assure reliability, the experiment was run three times and the temperature and distance of the UVB bulb to the plates was also kept constant at twenty centimeters. Additionally, the temperature measured from the plates during the UVB exposure was twenty nine degrees Celsius, extremely close to the temperature of the incubator, eliminating these confounding variables.
Results:
The objective of this experiment was to observe the effects of UVB light exposure on E. coli K12, and leading to various issues derived from that damage; for example, prohibited growth, cell death, and potential mutations.
The plates of E-Coli K12 were spread plated using a using a 10-4 dilution of E-Coli K12 and then immediately exposed to the reptile bulb for the amount of time allotted to each plate. This is either none (control), thirty minutes, sixty minutes, or ninety minutes of exposure under the reptile bulb, followed by immediate incubation. By conducting the trials in this manner, one can efficiently damage the bacteria and observe the following effect immediately after the bacteria grows. The term “sunlight” used in the discussion below refers to the light from the UVB bulb.
The results obtained are as follows:
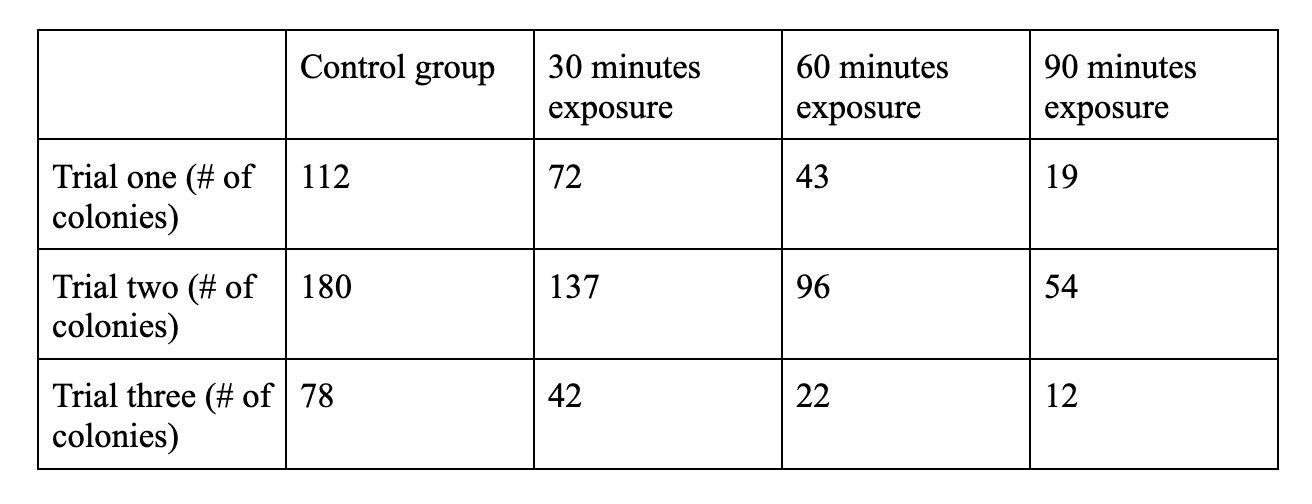
Figure 4: number of colonies remaining in each plate in the experiment, providing a clear distinction between the values of the control and each of the test groups
Due to the variety and differences between this data, despite clearly correlating a lengthened exposure time with more damage to the bacteria, and therefore less colonies, it is difficult to find a conclusive set of numbers from this data. However, due to various inconsistencies and variables involved in the growth of bacteria, along with various procedural errors, the results seem to only correlate within their own trial groups, and not among the various trials.
In order to remedy this, the data was converted into percent remaining of the control. This assumes the control as 100% of the potential growth, and the percentage of bacteria remaining is the bacteria still alive after UV exposure. The equation # of test group colonies / # of control colonies = % of control colonies remaining was used. For example, 72 test group colonies / 112 control group colonies = 64.3% of the control group.
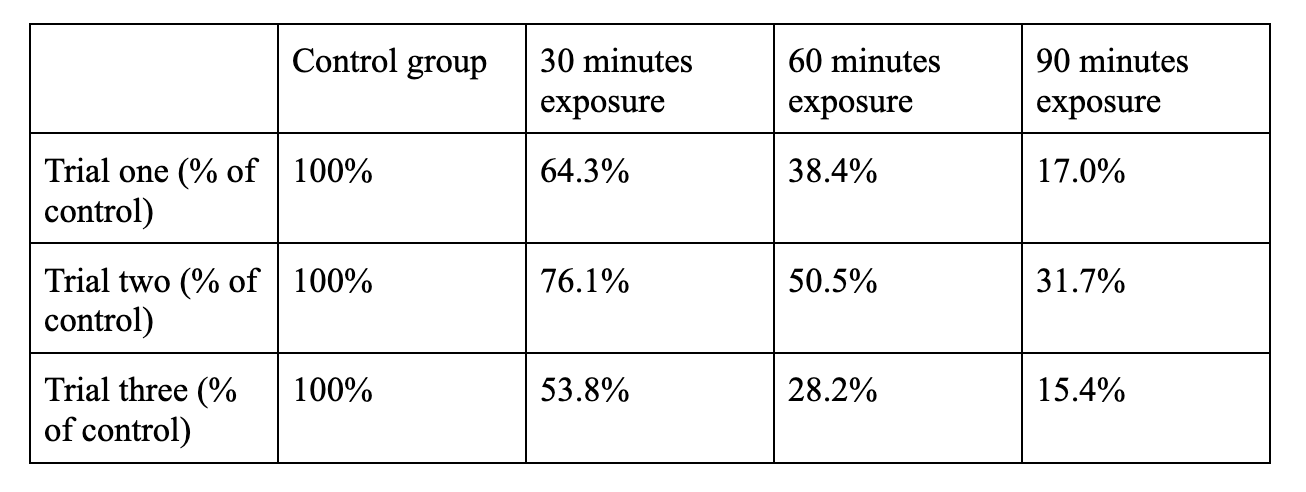
Figure 5: number of colonies transformed into percentage of control group remaining
With this data a chart was created to visualize the damage done by the UV light to the developing E-Coli.
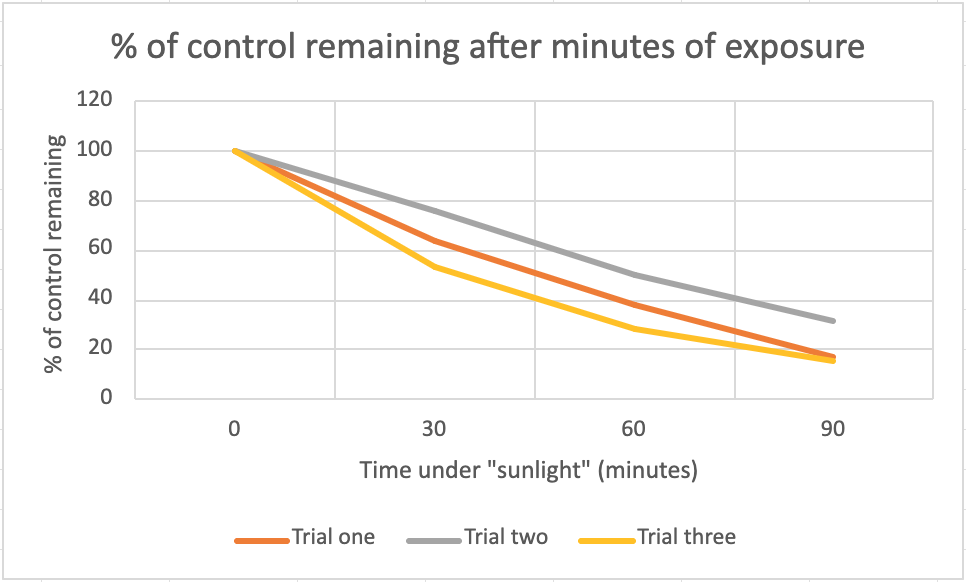
Figure 6: chart of data from figure 2
This chart illustrates the similarities between the levels of UV radiation that the bacteria were exposed to. It shows a similar downward sloping trend, meaning that each trial experiences a similar amount of damage in proportion to their respective control groups.
The results obtained from this experiment can be interpreted as the average of each of these relative percentages. After thirty minutes of exposure, 64.7% of the bacteria managed to grow, after an hour of exposure, 39.0% of the bacteria managed to grow, and after 90 minutes of exposure, 21.4% of the bacteria managed to grow. This strongly correlates a longer period of “sunlight” exposure leading to more damage to the bacteria, observed by noting the lower and lower percentage of the control groups remaining after longer periods of exposure.

Figure 7: This is an initial incomplete trial with an E-Coli concentration too high to effectively count colonies. Even without an accurate count, one can visually observe the density of bacteria colonies decline in relation to the increase in exposure time.
Discussion:
The results obtained from this experiment strongly correlate a longer period of UVB exposure with more damage to the bacteria, observed through a lower percentage of control colonies remaining after longer periods of exposure. These findings matched the initial hypothesis and the expected outcome. After 30, 60, and 90 minutes of exposure, only 64.7%, 39.0%, and 21.4% of colonies survived relative to their control groups, respectively. This is consistent with what was predicted going into this experiment, as UVB radiation is a proven mutagen, capable of inducing direct DNA damage through the formation of cyclobutane pyrimidine dimers and 6-4 photoproducts, both of which disrupt essential cellular pathways, and can ultimately lead to cell death or carcinogenesis.
It is important to note that this experiment was conducted in a controlled environment, with the bacteria being given a nutrient rich agar medium to grow and an optimal incubation temperature during, and immediately following their exposure period. The damage observed is therefore likely attributable to the UVB exposure itself, as major confounding variables such as temperature and exposure intensity were kept consistent throughout the experiment. If this experiment were conducted in an open or outdoor environment, the results would likely be significantly more variable, as many additional environmental factors would influence bacterial growth and survival beyond just UV exposure.
The results showed a similar trend across all three trials when converted to a percentage of the control, despite large variation in raw colony counts between trials. This variation is likely attributable to various inconsistencies in the experimental process. One inconsistency was the failure to immediately observe the bacteria after their twenty-four-hour growth period, often leaving the colonies in the fridge for one to two weeks before observation. Another inconsistency was the contamination of two of my trials, one of which had to be discarded, by a yellow bacterial contaminant. It seemed to be another organism whose colonies were small and yellow, covering the pale E-Coli, and not being affected by UVB light, completely hindering by ability to accurately count the E-Coli. The subsequent plate was also contaminated, however I could accurately count the E-Coli on this plate, as the density of the contaminant was lower.
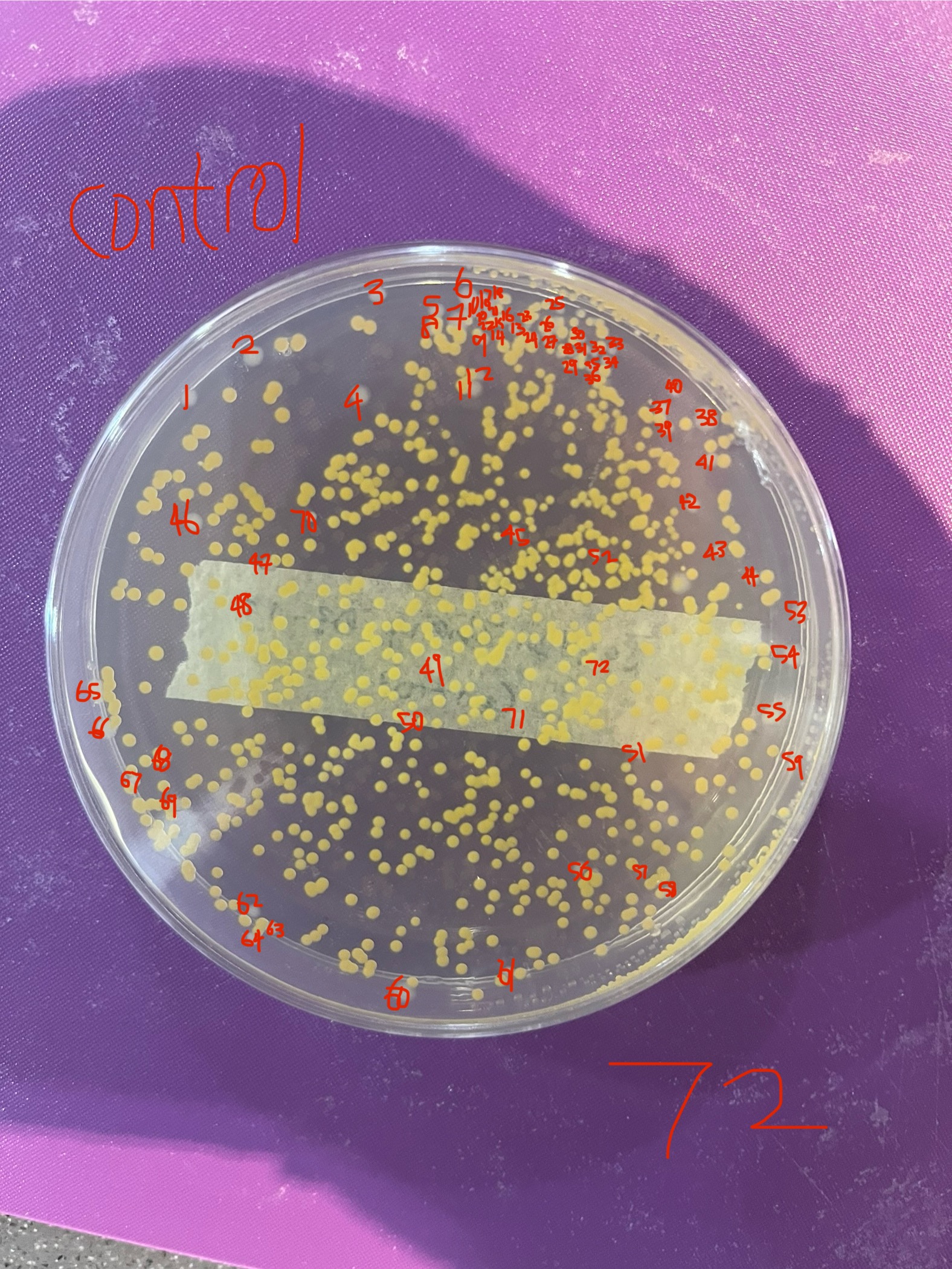
Figure 8: Pale white E-Coli K12 colonies covered by a yellow contaminant, with the E-Coli K12 still visible and countable.

Figure 9: The yellow contaminant density staying constant despite longer exposure times.
Converting the data to a percentage of the control was therefore a necessary step to meaningfully compare results across trials, and doing so revealed a similar downward sloping trend across all three experimental groups, further strengthening the correlation between UV exposure and bacterial damage despite roadblocks along the way.
There are also several limitations to this study worth acknowledging. The UVB reptile lamp used in this experiment is designed to mimic sunlight, but does not perfectly replicate the full spectrum of solar radiation, meaning the damage observed may not be entirely representative of the damage caused by natural sun exposure. Additionally, colony counts alone cannot distinguish between cells that were killed outright by UV exposure and cells that survived but sustained mutations, meaning the true extent of the DNA damage inflicted may be underrepresented by the data. Future studies could address this by examining surviving colonies for mutagenic changes and DNA damage.
Understanding how UV radiation damages bacterial DNA and inhibits growth is very relevant to our broader understanding of how UV radiation causes damage in human cells, ultimately contributing to skin cancer and carcinogenesis. The mechanisms of UV-induced DNA damage, such as the formation of CPDs and 6-4 photoproducts, are present across many forms of life, meaning the damage observed in E. coli K12 in this experiment is reflective of the same types of damage occurring in human skin cells upon UV exposure. Further research building on these findings could contribute to the development of photorepair technologies, better UV protection strategies, and a deeper understanding of the biological mechanisms behind skin cancer.
References:
Rastogi, R. P., Richa, N., Kumar, A., Tyagi, M. B., & Sinha, R. P. (2010). Molecular Mechanisms of Ultraviolet Radiation‐Induced DNA Damage and Repair. Journal of Nucleic Acids, 2010(1), 592980–592980. https://doi.org/10.4061/2010/592980
Kciuk, M., Marciniak, B., Mojzych, M., & Kontek, R. (2020). Focus on UV-Induced DNA Damage and Repair—Disease Relevance and Protective Strategies. International Journal of Molecular Sciences, 21(19), 7264. https://doi.org/10.3390/ijms21197264
Chang, M. S., Hartman, R. I., Trepanowski, N., Giovannucci, E. L., Nan, H., & Li, X. (2022). Cumulative Erythemal Ultraviolet Radiation and Risk of Cancer in 3 Large US Prospective Cohorts. American Journal of Epidemiology, 191(10), 1742–1752. https://doi.org/10.1093/aje/kwac101
Zhivagui, M., Hoda, A., Valenzuela, N., Yeh, Y.-Y., Dai, J., He, Y., Nandi, S. P., Burcak Otlu, Houten, B. V., & Alexandrov, L. B. (2023). DNA damage and somatic mutations in mammalian cells after irradiation with a nail polish dryer. Nature Communications, 14(1), 276–276. https://doi.org/10.1038/s41467-023-35876-8
Marie-Therese Leccia, Lebbe, C., Jean-Paul Claudel, Mridvika Narda, & Basset-Seguin, N. (2019). New Vision in Photoprotection and Photorepair. Dermatology and Therapy, 9(1), 103–115. https://doi.org/10.1007/s13555-019-0282-5
Ramírez-Gamboa, D., Díaz-Zamorano, A. L., Meléndez-Sánchez, E. R., Reyes-Pardo, H., Villaseñor-Zepeda, K. R., López-Arellanes, M. E., Sosa-Hernández, J. E., Coronado-Apodaca, K. G., Gámez-Méndez, A., Samson Afewerki, Hafiz, Parra-Saldivar, R., & Martínez-Ruiz, M. (2022). Photolyase Production and Current Applications: A Review. Molecules, 27(18), 5998–5998. https://doi.org/10.3390/molecules27185998
Gorini, F., Scala, G., Cooke, M. S., Majello, B., & Amente, S. (2021). Towards a comprehensive view of 8-oxo-7,8-dihydro-2’-deoxyguanosine: Highlighting the intertwined roles of DNA damage and epigenetics in genomic instability. DNA Repair, 97, 103027. https://doi.org/10.1016/j.dnarep.2020.103027
https://www.facebook.com/sag.micro. (2022, August 26). Spread Plate Method- Definition, Principle, Procedure, Uses. Microbe Notes. https://microbenotes.com/spread-plate-technique/