Sofia Nacho — Year 2, Life Science
Introduction
In recent years, studies have increasingly emphasized the impact that the gut’s microbiome has on animals, including humans. This highlights the importance of understanding natural methods to support gut health, particularly during periods of microbial imbalance (dysbiosis)
Surveys from recent decades rank Canada among the highest in incidence rates of inflammatory bowel disease (IBD) and irritable bowel syndrome (IBS). IBD involves chronic inflammation of the digestive tract due to immune system dysfunction, whereas IBS is associated with dietary triggers and is primarily managed through symptom control. Whereas, IBS involves certain trigger foods, so diet is used as a primary treatment for symptom control. While IBD is more severe, IBS is significantly more common, affecting approximately 18% of Canadians compared to roughly 1% for IBD.
The gut’s microbiome is a complex, living ecosystem that is very diverse, containing a significant population of viruses, fungi, archaea, but mostly bacteria (Ravichandra Vemuri et al., 2020). For this reason, bacteria (and the types of bacteria) are especially important to human health, and they are known as the gut microbiota. The microbiota is crucial for the body’s digestion and metabolism, producing many essential nutrients. Certain food, especially dietary fibers, support the growth of healthy gut bacteria by reducing oxygen levels when fermented by the gut’s bacteria. By lowering oxygen levels, redox potential is also lowered, meaning it makes the way electrons are taken or given by the chemical molecules in the gut beneficial for fermenting bacteria and harmful for oxygen-loving bacteria (Ravichandra Vemuri et al., 2020). Although oxygen is essential for human survival, a healthy gut environment is typically characterized by low oxygen levels that support anaerobic microbial populations
According to a study done by Rongying Xu et al. (2024) pectin (a polysaccharide found in the cell walls of plants) was found to help the gut microbiome recover faster and function better after it was supplemented into rats that were previously suffering from dysbiosis through antibiotics. In addition to aiding recovery, pectin strengthens the gut barrier, prevents harmful microbial invasion, and reduces intestinal inflammation, also strengthening the gut barrier. This prevents harmful microbes from entering, and ultimately reducing intestinal inflammation (Sammra Maqsood et al., 2025). Pectin is also a prebiotic, meaning it supports the probiotics that can still be found in a gut suffering from dysbiosis.
Since Pectin is found in the cell walls of all plants (Monika Szymańska-Chargot et al.,2014), it plays a crucial role in providing structure and yet flexibility to a plant’s tissue. Apple Pomace is especially abundant in this fiber compound, making apples an excellent resource when striving for a more nutritionally beneficial diet. When it comes to extracting pectin, Hydrolysis is a conventional method for a Functional outcome enabling efficient industrial extraction of pectin while increasing extraction yield (how much pectin can be separated) by accelerating cell wall disruption and mass transfer. According to an applied sciences journal by Anissa Belkheiri et al., (2021) on extraction of pectin from plant-based products, if low pH and high temperatures are maintained for long periods of time, pectin release can be effectively promoted through hydrolysis. However, their study also highlights the fact that this more traditional method could result in pectin degradation and its overall resulting quality. Degradation of pectin means that viscosity (the pectin’s natural sticky thickness) is reduced as the molecules are broken into shorter chains which lowers their molecular weight. For this reason, the paper deliberately avoids hydrolysis since for its area of research, it is crucial that the natural structure of pectin is preserved.
Even so, if the objective is to test the effect that pectin’s prebiotic nature has on different probiotics, the reduced chain length and esterification would still yield accurate results. A study on pectin hydrolysates by Ho, Yen-Yi et al., (2017) showed that the smaller molecular‑weight fractions produced by hydrolysis significantly stimulated the growth, fermentation, and survival of probiotic strains. Essentially, when larger molecules are broken down into smaller pieces, they help bacteria work better, grow more efficiently, and stay alive for longer. In their study, pectin was still able to act as a prebiotic base material that supported the growth of probiotics or beneficial gut bacteria. This shows that the high-molecular-weight pectin is not crucial for result yielding fermentation studies. Therefore, even partially degraded pectin can serve as a prebiotic substrate, meaning it is indigestible and can only be fermented by the gut’s microbes.
The importance of prebiotics is highlighted when they work alongside probiotics. Probiotics are living microorganisms that naturally balance the microbiome by boosting immunity. They help with digestion and taking space and nutrients from harmful bacteria (Sammra Maqsood et al., 2025). As fibers, prebiotics fuel and sustain the guts existing bacteria, while probiotics themselves are beneficial microbes. In this way, both provide the most benefits when the other is present, prebiotics as healthy fuel and probiotics as healthy things to get fueled (Paulina Markowiak and Katarzyna Slizewska, 2017)
One recent and fast expanding area of research is the relationship that the gut’s community of microorganisms has on mental health. New studies are uncovering how an organism’s tendency to be depressed, their ability to regulate stress chemicals, and their quality of cognitive function are dependent on the gut’s state. Will Houston (2025) posted a scientific research article revealing how adulthood depression and emotional regulation is likely related to their microbiomes state in early childhood. This suggests that the gut plays a key role in the development of emotional brain circuits. It is common knowledge that depression rates have gone up in Canadians in recent decades and concurrently, the average citizen’s diet has become more westernized tending to rely on fast food with less of an emphasis on fiber intake.
Many studies have investigated the positive individual impacts of the prebiotic pectin on probiotics (Rongying Xu et al. 2024) but not specifically extracted from Canadian native ambrosia apples. Their widespread availability, high nutritional value and general appeal make them one of the most popular fruits in hospitals, schools, and shelters.
This study aims to investigate how apple-derived pectin extracts influence the growth and metabolic activity of probiotic bacteria across different food-based environments. Just as a snack, apples can be easily implemented into any diet, and their easy access makes distribution even more efficient. By denoting the most bacteria supporting combination of apple pectin and other probiotics, the ground can be set for reaping as much of the benefits that such a widespread food can offer.
Materials and Methods
Materials
Apples were Ambrosia and on average weighed 180g. The apples were bought fresh from a local market and cultivated in British Columbia during 2026. The apples were chopped and their seeds were removed but the peel was included as it contains much of the apple’s pectin. The apple mash blended from these apples was hydrolysed with a commercial enzymatic preparation: Elo’s Premium Pectic Enzyme (100g).
Apple Mash Preparation
Apple pectic oligosaccharides (POS) were prepared by controlled enzymatic hydrolysis of apple mash. Fresh Ambrosia apples were washed thoroughly, cored to remove seeds, and blended whole (including peels) with distilled water at a 1:1 (w/v) ratio to ensure retention of pectin-rich peel material.
The resulting mash was heated to 85–90°C for 20 minutes with occasional gentle stirring to dissolve pectin, weaken cell wall structure, and improve receptivity to the enzymatic hydrolysis. This also ensures that apple mash does not burn at the bottom of the container.
Sodium Acetate Buffer
During the heat treatment, a 0.05 M sodium acetate buffer (pH ~5.) was prepared by dissolving 4g of sodium acetate in 100ml distilled water. This buffering condition was chosen to approximate the optimal pH range for pectinase activity in imitation of research done by Milos V. Nikolic and Ljiljana Mojovic (2006). Before the buffer is added, the apple mixture should have a pH around 3.5. when added, pH of apple solution should be 4.5.
Hydrolysis
After heating mash, the mash was cooled to 45–50°C. Sodium acetate buffer was added, and two tablespoons (30ml) of pectinase was introduced at twice the standard recommended dosage to compensate for the shorter hydrolysis period. Enzymatic hydrolysis happened at 45–50°C for 60–75 minutes under gentle mixing as before. The hydrolysis was ended by heating the mixture to 95–100°C for 10 minutes to fully denature the enzyme.
Clarification
The hydrolysate was clarified by filtration through a coffee filter (cheese cloth works), and the liquid was collected. The resulting extract consisted of a liquid apple hydrolysate enriched in pectic oligosaccharides (degree of polymerization approximately 2–10) and residual soluble sugars.
Plating
For plating, there were three experimental groups, three food control groups, and one pectin control group. Standard agar plates were used with a 90 mm diameter. The experimental groups had 2 ml of the pectin rich apple solution and 2 ml of one of the food growth mediums (either yogurt, kefir, or milk). The control groups each had 2ml of only one type of growth medium (yogurt, kefir, milk, and apple pectin solution). This ensures that any change in bacterial growth between food cultures is due to the added presence of pectin. Three trials were conducted for each condition, and they were each incubated for a week at 50 °C.
Results
This research was conducted to measure the microbial growth with pectin acting as a prebiotic to stimulate probiotic growth across different possible food pairings (milk, yogurt, and kefir). Pectic oligosaccharides (POS) were successfully derived from the apple pectin’s hydrolysis.
To verify the results, three trials were conducted, all of which produced consistent outcomes, three official trials were made, which all displayed consistent results. All the groups with isolated probiotics as well as pectin by itself, displayed no signs of microbial growth.
The plates with both apple pectin extract and the probiotic food sources had the most microbial growth compared to the control plates containing only the probiotic food sources. Increased microbial growth in the experimental groups can be explained by the addition of pectic oligosaccharides (POS). Once in oligosaccharide form, these compounds can be transported across bacterial cell membranes through carbohydrate transport systems. In many types of probiotics, these substrates are turned into intermediates that enter central carbon metabolism, leading to the generation of ATP which is needed for cell replication.
The reduced structural complexity of oligosaccharides (compared to intact polysaccharides) significantly increased the number of colonies. This is because shorter carbohydrate chains require fewer enzymatic steps to metabolize, allowing bacteria to spend more energy toward growth and reproduction rather than breaking down sugars. This provides a clear explanation for why the addition of hydrolyzed pectin led to greater microbial growth. (Flint, Harry J. et al. 2012).
Visual Observations
Kefir and Pectin:
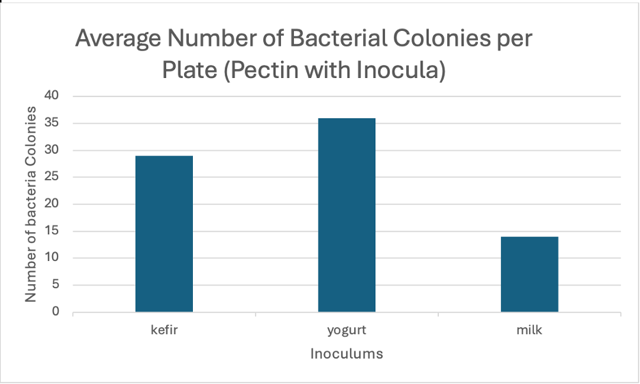
Kefir showed the second most colony formation after pectin.
Yogurt and Pectin:
With the addition of pectin extract, yogurt showed the most density and quantity of colony formation out of the three growth tested media.
Milk and Pectin:
Milk was the only inoculum, which was not fermented, resulting in it not containing fewer colonies compared to yogurt or kefir.
Data
| Trials: | Kefir+Pectin | Yogurt+Pectin | Milk+Pectin |
| #1 | 27 | 36 | 12 |
| #2 | 31 | 38 | 17 |
| #3 | 30 | 35 | 13 |
| Average | 29 | 36 | 14 |
Figure 1: raw data from all official trials and the average used in graph



Discussion
Initially, apple pomace was used for agar plating; however, this method created difficulties in colony counting due to its uneven texture, but this method raised complications for plate reading because it was difficult to count the colonies due to the mush’s uneven structure. For the results, the liquid component of the apple mash was used instead since it created a flatter surface for colonies to grow and be observed. This liquid apple solution will likely still show pectin’s effect on different food mediums since hydrolysis has allowed the pectin to separate. This allows probiotic bacteria to get an instant energy source without requiring more extracellular breakdown
Using resulting liquid instead of pomace also provides greater clarification.
It was hypothesized that kefir would exhibit the highest bacterial growth due to its typically higher Lactobacillus content have the most bacterial growth since it typically has a greater Lactobacillus (a genus of bacteria that is commonly found in fermented foods and considered beneficial for the human gut) population than yogurt. In this case, Greek Yogurt was specifically used which is known to be denser in bacteria per ml, creating more colonies than a basic yogurt brand would (Espinosa-Andrews et al., 2025). This supports faster bacterial reproduction, explaining the higher colony densities observed on plates containing both pectin and probiotic sources.
As expected, the plate with pectin and milk had the fewest number of countable colonies. It was assumed that this was because milk was the only inoculum, which was not a fermented product, resulting in it not containing a significant number of probiotics.
Limitations
This study is limited by a small sample size (n = 3) and a lack of quantitative precise measurement. There is also an absence of chemical verification of pectic oligosaccharide production, with only general colony formation being observed. For this reason, the species of bacteria that were able to form colonies could not be identified. Also, microbial populations in the tested food sources were not controlled, and incubation at 50 °C may not have been optimal for probiotic growth. This may have inhibited or reduced the growth of certain probiotic species.that might otherwise have prospered in the gut. As for filtering methods for the pectin after hydrolysis, it increased the potential contamination, ultimately reducing reliability. Finally, this study did not distinguish between the effects of oligosaccharides and simple sugars released during hydrolysis, limiting the conclusions regarding prebiotic activity.
Future research
The resulting plates in the investigation were incubated at 50 °C, but lower temperatures may be more optimal for bacterial growth in future experiments. Incubating the plates at 37 °C (Vemuri, R. et al. 2020), which is a more standard temperature, may reveal more accurate bacteria colony diversity and quantity. Future investigations could also test for the possible effects that temperature has on these specific pectin and probiotic groups. This could provide insight into the thermal resilience of microbial populations in the presence of pectin with the support of pectin, revealing the protective effects of oligosaccharides.
References
Ravichandra Vemuri et al., (2020) Beyond Just Bacteria: Functional Biomes in the Gut Ecosystem Including Virome, Mycobiome, Archaeome and Helminth. https://pubmed.ncbi.nlm.nih.gov/32231141/
Vemuri, R., Shankar, E. M., Chieppa, M., Eri, R., & Kavanagh, K. (2020). Beyond Just Bacteria: Functional Biomes in the Gut Ecosystem Including Virome, Mycobiome, Archaeome and Helminths. Microorganisms, 8(4), 483. https://doi.org/10.3390/microorganisms8040483
Rongying Xu et al. (2024). Pectin supplementation accelerates post-antibiotic gut microbiome reconstitution orchestrated with reduced gut redox potential https://pmc.ncbi.nlm.nih.gov/articles/PMC11203915/#abstract1
Xu, R., Feng, N., Li, Q., Wang, H., Li, L., Feng, X., Su, Y., & Zhu, W. (2024). Pectin supplementation accelerates post-antibiotic gut microbiome reconstitution orchestrated with reduced gut redox potential. The ISME journal, 18(1), wrae101. https://doi.org/10.1093/ismejo/wrae101
Sammra Maqsood et al., (2025) Fruit‐Based Diet and Gut Health: A Review. https://pmc.ncbi.nlm.nih.gov/articles/PMC12041667/#fsn370159-sec-0004
Maqsood, S., Arshad, M. T., Ikram, A., & Gnedeka, K. T. (2025). Fruit-Based Diet and Gut Health: A Review. Food science & nutrition, 13(5), e70159. https://doi.org/10.1002/fsn3.70159
Monika Szymańska-Chargot et al., (2014) Raman imaging of changes in the polysaccharides distribution in the cell wall during apple fruit development and senescence. https://pmc.ncbi.nlm.nih.gov/articles/PMC4819746/
Szymańska-Chargot, M., Chylińska, M., Pieczywek, P. M., Rösch, P., Schmitt, M., Popp, J., & Zdunek, A. (2016). Raman imaging of changes in the polysaccharides distribution in the cell wall during apple fruit development and senescence. Planta, 243(4), 935–945. https://doi.org/10.1007/s00425-015-2456-4
Anissa Belkheiri et al., (2021) Extraction, Characterization, and Applications of Pectins from Plant By-Produccts. https://www.mdpi.com/2076-3417/11/14/6596
Belkheiri, A., Forouhar, A., Ursu, A. V., Dubessay, P., Pierre, G., Delattre, C., Djelveh, G., Abdelkafi, S., Hamdami, N., & Michaud, P. (2021). Extraction, Characterization, and Applications of Pectins from Plant By-Products. Applied Sciences, 11(14), 6596. https://doi.org/10.3390/app11146596
Ho, Yen-Yi et al., (2017) Evaluation of the prebiotic effects of citrus pectin hydrolysate. https://pubmed.ncbi.nlm.nih.gov/28911641/
Ho, Y. Y., Lin, C. M., & Wu, M. C. (2017). Evaluation of the prebiotic effects of citrus pectin hydrolysate. Journal of food and drug analysis, 25(3), 550–558. https://doi.org/10.1016/j.jfda.2016.11.014
Paulina Markowiak and Katarzyna Slizewska, (2017) Effects of Probiotics, Prebiotics, and Synbiotics on Human Health. https://www.mdpi.com/2072-6643/9/9/1021?utm_campaign=CHD_%25D9%2585%25D8%25B1%25D8%25A7%25D8%25AC%25D8%25B9%25D8%25A7%25D8%25AA-%25D8%25A5%25D9%2586%25D8%25B2%25D9%258A%25D9%2585%25D9%2585%25D8%25AF
Markowiak, P., & Śliżewska, K. (2017). Effects of Probiotics, Prebiotics, and Synbiotics on Human Health. Nutrients, 9(9), 1021. https://doi.org/10.3390/nu9091021
Houston. W. (2025) Babies’ Gut Bacteria May Influence Future Emotional Health. https://www.uclahealth.org/news/release/babies-gut-bacteria-may-influence-future-emotional-health#:~:text=A%20child’s%20early%20gut%20microbiome,said%20study%20senior%20author%20Dr.
Schriger, D. L., Arora, S., & Altman, D. G. (2006). The content of medical journal instructions for authors. Annals of Emergency Medicine, 48(6), 743–749.e1–4. https://doi.org/10.1016/j.annemergmed.2006.03.028
Vemuri, R., Shankar, E. M., Chieppa, M., Eri, R., & Kavanagh, K. (2020). Beyond Just Bacteria: Functional Biomes in the Gut Ecosystem Including Virome, Mycobiome, Archaeome and Helminths. Microorganisms, 8(4), 483. https://doi.org/10.3390/microorganisms8040483
Flint, H. J., Scott, K. P., Duncan, S. H., Louis, P., & Forano, E. (2012). Microbial degradation of complex carbohydrates in the gut. Gut microbes, 3(4), 289–306. https://doi.org/10.4161/gmic.19897
Espinosa-Andrews, H., Haro-González, J. N., Barbosa-Nuñez, J. A., Aguirre-Mandujano, E., & Lobato Calleros, C. (2025). Physicochemical and rheological characteristics of commercial Greek-style yogurt enriched with Polygonum cuspidatum roots or the P. cuspidatum β-cyclodextrin inclusion complex. Food Research International, 203, 115854. https://doi.org/10.1016/j.foodres.2025.115854v