Sophia Liu — Year 2, Life Science
Abstract
Magnesium-containing minerals are found to exist on Mars, and they play an important role in the chemistry of Martian surfaces and are an essential element for bacterial growth. Therefore, it is necessary to examine the tolerance of microbes to high magnesium environments, especially when high concentrations may affect ionic balances and the survival of organisms. This study aims to examine the effects of increasing magnesium sulphate concentrations on the colony formation of Escherichia coli K-12 through exposing bacterial cultures to different initial concentrations of magnesium sulphate before being transferred to a higher, uniform level of magnesium sulphate concentration. The observations of this study suggest that as the concentration of magnesium is increased, the number of bacterial colonies decreases.
Introduction
In the scientific world of biochemistry and natural sciences, the study and understanding of how life responds to extreme environments is an important step in advancing research, especially in fields of microbiology and astrobiology. On Earth, living microorganisms have been identified in nearly every environment, even those with high salinity, extreme temperatures, and unusual chemical compositions. Therefore, due to the versatility and high reproduction rates of bacteria, these organisms have been commonly used as models to study biological responses to environmental stress. When looking at extraterrestrial environments such as Mars, these studies are especially relevant as extraterrestrial surface environments significantly differ from those on Earth. One major difference is soil chemistry, which may include high concentrations of salt and metal ions, such as magnesium-containing compounds (Scheller et al., 2021).
Magnesium-containing minerals have been identified on Mars and are believed to influence the surface chemistry of the planet (Crisler et al., 2012; Scheller et al., 2021). Scheller et al. (2021) discusses the implications of how the formation of magnesium carbonates in Martian environments may affect planetary habitability. While this work is more closely linked to geochemistry, it is essential to note that its implications are valid for researching effects on living organisms as it shows that magnesium-containing compounds are likely found in considerable quantities in extraterrestrial environments. Hence, if life were to survive in such conditions, it would have to be able to withstand high concentrations of magnesium.
Microorganisms are often used to test the above limits due to their relative simplicity and ability to survive in a wide range of habitats. In a study by Crisler et al. (2012) examining bacterial growth in high concentrations of magnesium sulphate in a stimulation of Martian soil, some bacteria are found to be capable of surviving and growing in high magnesium sulphate concentrations. This study shows that high salinity conditions are not necessarily a barrier to microbial life and proves the value of using bacteria to test these limits.
Escherichia coli (E. coli) is one of the most widely used model organisms among the bacterial population. E. coli grows rapidly and can be cultured using standard laboratory protocols, which makes it the preferred organism of study to test the effects of environmental variables. One of the earliest studies on the effects of magnesium on the growth of E. coli was conducted by Lusk et al. (1968). This study set the stage for human understanding of the importance of magnesium in the growth of bacteria by demonstrating that normal bacterial growth is driven by the presence of magnesium.
However, the biological importance of magnesium is not limited to growth alone. Magnesium ions (Mg²⁺) play essential roles in many cellular processes and functions. Walker (1994) explored the role of magnesium ions in biotechnology and cellular biology, emphasizing the role of magnesium ions as a cofactor for enzymes, as well as their contribution to the stabilization of cellular membranes and the maintenance of nucleic acids. Furthermore, the presence of magnesium is crucial for ribosomal stability, which is vital since ribosomes are directly involved in protein synthesis, which is the foundation for the basic survival of all organisms. Without an adequate amount of magnesium, ribosomes will not function sustainably, and cellular division will be significantly affected. Conversely, an excess amount of magnesium can cause stress on an organism due to an imbalance of ions within the cells.
Due to both the importance and potential stress caused by magnesium, bacteria must be able to regulate the concentration of magnesium inside their cells. This process is known as magnesium homeostasis, and is a way to help the cell adapt to changes in the magnesium concentration of the environment. The role of magnesium homeostasis in bacterial adaptation to osmotic stress was studied by Wendel et al. (2022). The results of the study supported the idea that the regulation of magnesium is crucial for the adaptation and further development of bacteria after being exposed to environmental stressors.
More recent studies have specifically addressed the response of E. coli to increasing magnesium sulphate concentrations. In the comprehensive study by Nepal and Kumar (2020), the effects of increasing E. coli exposure to magnesium were investigated in terms of how it affected bacterial growth, cell division, and gene expression. This research article showed that E. coli colonies were able to grow normally in increasing concentrations of magnesium sulphate up to 1.25 M before becoming significantly smaller in size.
The present experiment studies how increasing magnesium sulphate concentrations in solid LB growth media may affect the colony formation and growth of E. coli. Bacterial cultures of different initial concentrations are compared with cultures transferred to high concentrations of magnesium sulphate to assess growth under increased stress, and by examining the changes in colony density, color, and comparisons between colonies “buffered” in an extra week at the same concentration, this study will provide insight into the bacterial tolerance response at high-magnesium environments chemically relevant to extraterrestrial surfaces.
Materials and Methods
2.0M Magnesium Sulphate Stock Solution
A 100 mL beaker was weighed and recorded. About 24.078 g of BioShop Canada MgSO4 anhydrous reagent grade was added to the beaker and then recorded. A 200 mL beaker was placed on a stir plate with a magnetic stirring bar, to which about 80 mL of distilled water was added. The water was stirred on medium-high while the magnesium sulphate powder was slowly added to the solution. The beaker was mixed thoroughly to make sure the solution was fully dissolved. After two minutes of stirring, more distilled water was added to reach the 100 mL mark, before being left alone to stir on medium-high for five more minutes.

Figure 1: Set-up of how the 2.0M magnesium sulphate stock solution was made
Pouring Magnesium-Infused Plates
The C1V1 = C2V2 equation was used to calculate the amount of 2.0M stock needed to make a 25 mL growth medium mixture infused with the desired concentration of magnesium. The calculated amount of magnesium stock was measured in a 10 mL plastic graduated cylinder before it was transferred to a 50 mL glass graduated cylinder. Heated agar was poured into the graduated cylinder to the 25 mL mark. The graduated cylinder was then lightly swirled around before being poured into a sterile Petri dish labeled with the concentration, date, and initials of the experimenter.
Creating E.coli Bacterial Suspensions
A 10-5 serial dilution was used to create bacterial suspensions. A microtube rack was prepared and six microcentrifuge tubes was placed on it, labeled 1, 10-1 , 10-2 , 10-3 , 10-4 , 10-5 respectfully. A 0.9% NaCl solution was prepared to use in the dilution; 1500 mL of saline solution was put into the “1” tube, before following amounts of saline solution calculated as 90% of the desired amount of total solution was put into each of the other tubes respectfully. A 1000 µL and 200 µL micropipette will be needed to complete this process, as well as a waste beaker with bleach. Bacterial samples from a week prior will be used to make the bacterial suspension, and an inoculation hoop was used to touch three colonies before being swirled in the saline solution and vortexed for around 10 seconds. Then the process of transferring the desired amount of the previous suspension to the next one and vortexed was repeated until the serial transfer reached the last tube labeled 10-5.
Plating Suspensions
The bacterial suspension was plated onto the prepared plates using the spread plating technique. 100 µL of 10-5 bacterial suspension was pipetted onto the agar and spread plated using an L-spreader. Once all agar suspensions were poured, the plates were left to set for a minute before being inverted, plastic wrapped, taped together and incubated at 37 degrees Celsius for 24 hours.
Table 1: Table of which concentration each plate was transferred to by the end of the experiment.
| Original Plate | Transferred To |
| 0.15M, 0.25M, 0.35M | 0.15M Buffer, 0.25M Buffer, 0.35M Buffer |
| 0.15M, 0.25M, 0.35M | All to a concentration of 0.50M |
| 0.15M Buffer, 0.25M Buffer, 0.35M Buffer | All to a concentration of 0.50M |
Counting Colonies
At the start of each session, pictures of each plate was taken with both the front and back side. Data from these photos were analyzed with the editing function in an iPad’s built-in photo app by doting the colonies with red dots, then marking each 100 dots counted and summing the numbers to get the total amount of colonies. Plates with slight overgrowth were counted using the quadrant counting method, where the plate was split into four sections and counted one of them before multiplying across all four sections.
Concentrations Tested
Table 2: Magnesium Concentrations Tested
| Date | Magnesium Sulphate Concentration Tested |
| Jan 27, 2026 | 0M |
| Feb 3 | 2x Dilution, 3x Dilution |
| Feb 11 | 0.50M |
| Feb 17 | 0.10M |
| Feb 24 | 0.15M, 0.25M, 0.35M |
| Mar 3 | 0.15M Buffer, 0.25M Buffer, 0.35M Buffer, 0.15M0.50M, 0.25M0.50M, 0.35M0.50M |
| Mar 10 | 0.15M Buffer0.50M, 0.25M Buffer0.50M, 0.35M Buffer 0.50M |
Introducing Buffer Weeks
The key differentiation between this study with others is the introduction of “buffer weeks”, in which a strain of bacteria is exposed to one extra week of growth under the same magnesium concentration before being transferred to the increased magnesium concentration. Buffer weeks was introduced in the final weeks of the experiment.
Results
The results for this experiment varied greatly. For the first two weeks, E. coli colonies were not dilute enough to form CFU groups, therefore the data gathered were discarded.
During Week 3, two control groups and two 0.50M groups were grown but some yellow spots were present, indicating contamination. The same four plates were placed in the incubator for another 24 hours to test whether the small colonies from the 0.50M group would grow better, while new colonies at 0.10M and a control were grown.
By the fourth week, the four plates from week 3 seemed to have colonies grown bigger yet the yellow spots seemed to have spread, especially amongst the control groups which had yellow lines running along the edges of the agar plate. The new control and two 0.10M plates had completely yellow colonies, which was thought to be a spread of contamination since the plates were grown from the samples already with yellow spots from a week prior, so during week five the results from week four was discarded and four new plates from completely new bacterial was grown; one as a control with zero magnesium with the other three at 0.15M, 0.25M and 0.35M respectively.
During week six these plates were found to have grown completely yellow colonies as well which was strange considering they were “new” bacteria, and other plates from other experiments also had plates with completely yellow colonies. Hence, the results were kept, and seven more plates were plated. The experimental groups at 0.15M, 0.25M and 0.35M were both replicated at the same concentration and transferred to a 0.50M concentration at the same time. Therefore, each experimental group had two new plates plated, one to test the idea of a “buffer week” to see if bacteria exposed to the same concentration of stress could have better tolerance in the long run and the other to test the tolerance of bacteria to stress change from a short jump to a long one. During week six, these six new plates were plated along with a control group from the four plates made during week five.
Table 3: CFUs from all experimental plates
| Date | Countable Colonies (in CFU) |
| Jan 27, 2026 | N/A (Above CFU Count) |
| Feb 3 | N/A |
| Feb 11 | N/A |
| Feb 17 | N/A |
| Feb 24 | N/A (Above CFU Count) |
| Mar 3 | Control: 613. 0.15M Buffer: 404. 0.25M Buffer: 447. 0.35M Buffer: 364. 0.15M-0.5M: 370. 0.25M-0.50M: 245. 0.35M-0.50M: 351. |
| Mar 10 | Control: 265. 0.15MBuffer→ 0.50M: 244. 0.25MBuffer→ 0.50M: 1050. 0.35MBuffer→ 0.50M: 724. |
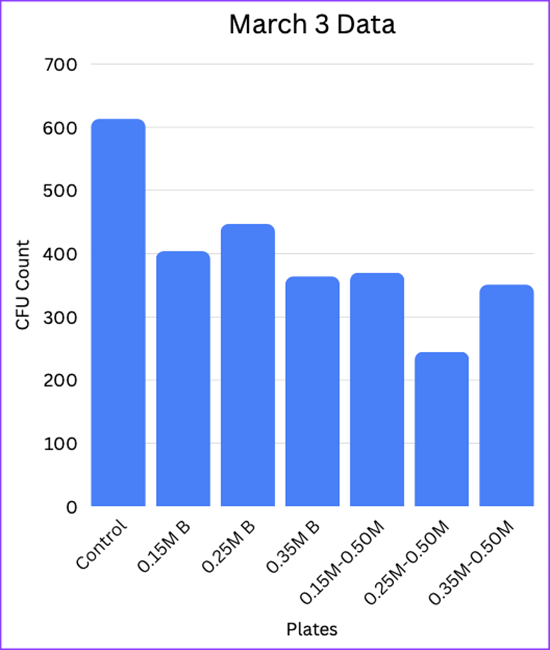
Figure 2: Graph of March 4th data

Figure 3: Graph of March 10th data
All the colonies were hand-counted using the photo app and an apple pencil to dot the colonies. As such, the transfer of buffer concentrations to 0.50M from Mar 10th were all estimates using the quadrant counting method. All other plates were counted entirely.
Looking at the data gathered from the experiment, it is clear that the introduction of a buffer week helped with the survival rates of bacterial colonies once introduced to a higher magnesium concentration. Though the plate transferred to 0.50M from the 0.15M Buffer had errors in colony formation due to an uneven agar surface, and the plate from 0.25M had a slight overgrowth into more than a thousand colonies, we can still look at the difference between transferring the 0.35M concentration directly to 0.50M compared to the buffer plate that had the same colonies plated at 0.35M for one more time before being transferred to 0.50M. According to the gathered data, colony growth went from 351 CFU in the 0.35M plate to 724 CFU in the 0.35M Buffer plate, more than doubling in amount of colony growth and survival. The 0.25M plates show a greater difference, going from 245 CFU to 1050 CFU, which is more than four times the amount of survivability. Therefore, it is credible to say that the introduction of a buffer week significantly aided the survivability of E. coli colonies when being transferred to a higher magnesium concentration.
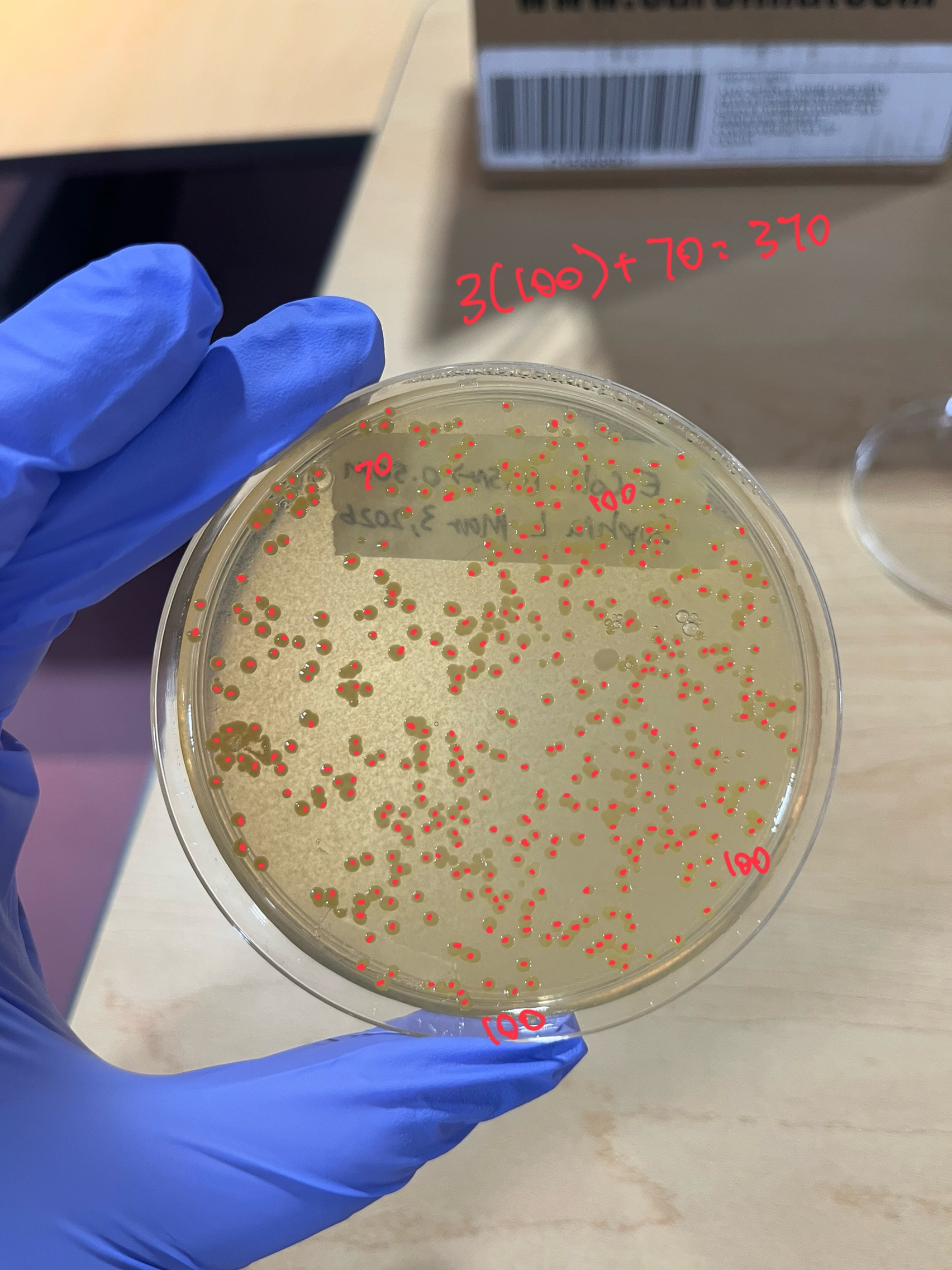
Figure 4: March 4th 0.15M → 0.50M plate, which was counted by the whole plate
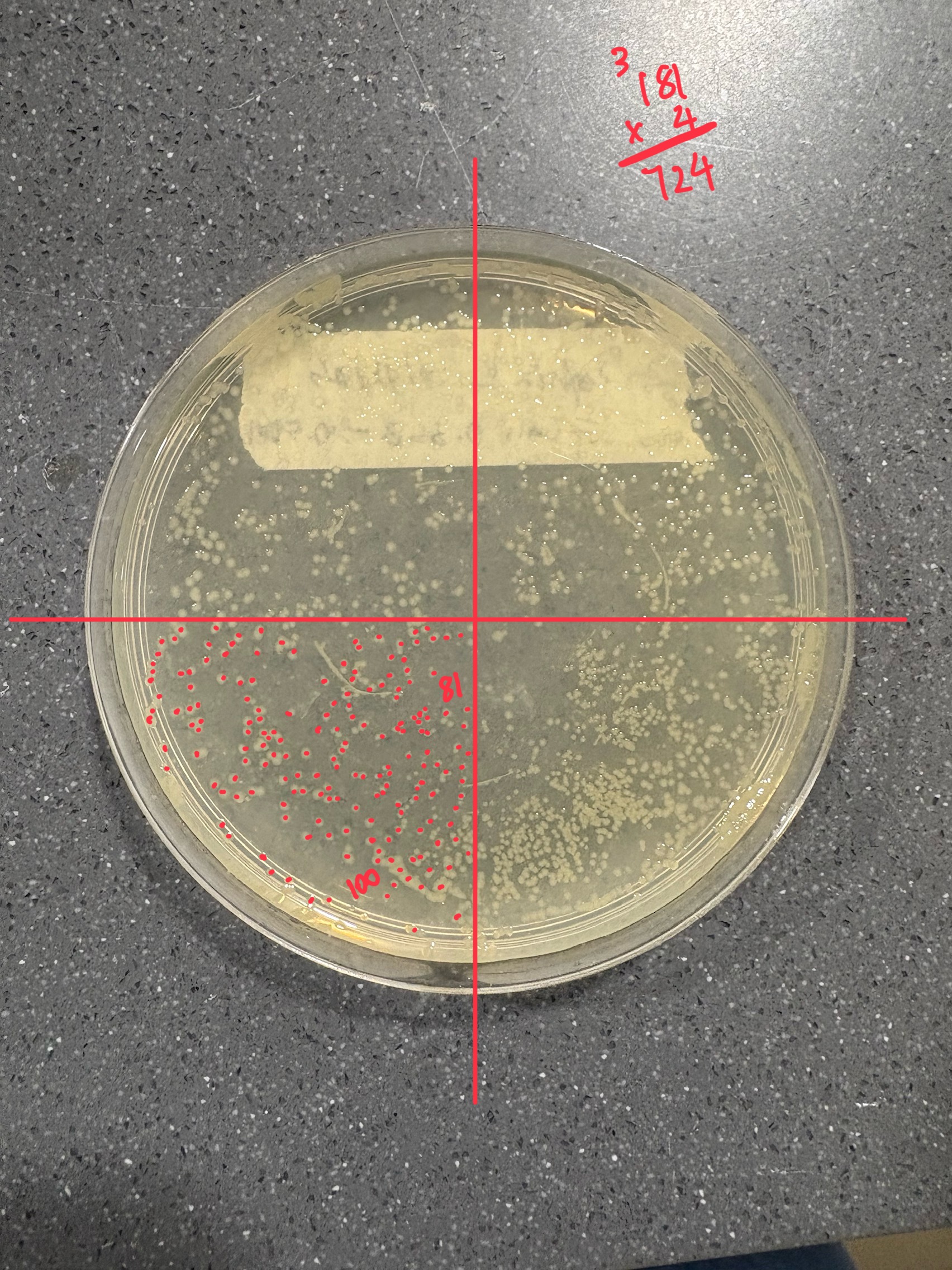
Figure 5: March 10th 0.35MBuffer → 0.50M plate, which was counted using the quadrant counting method
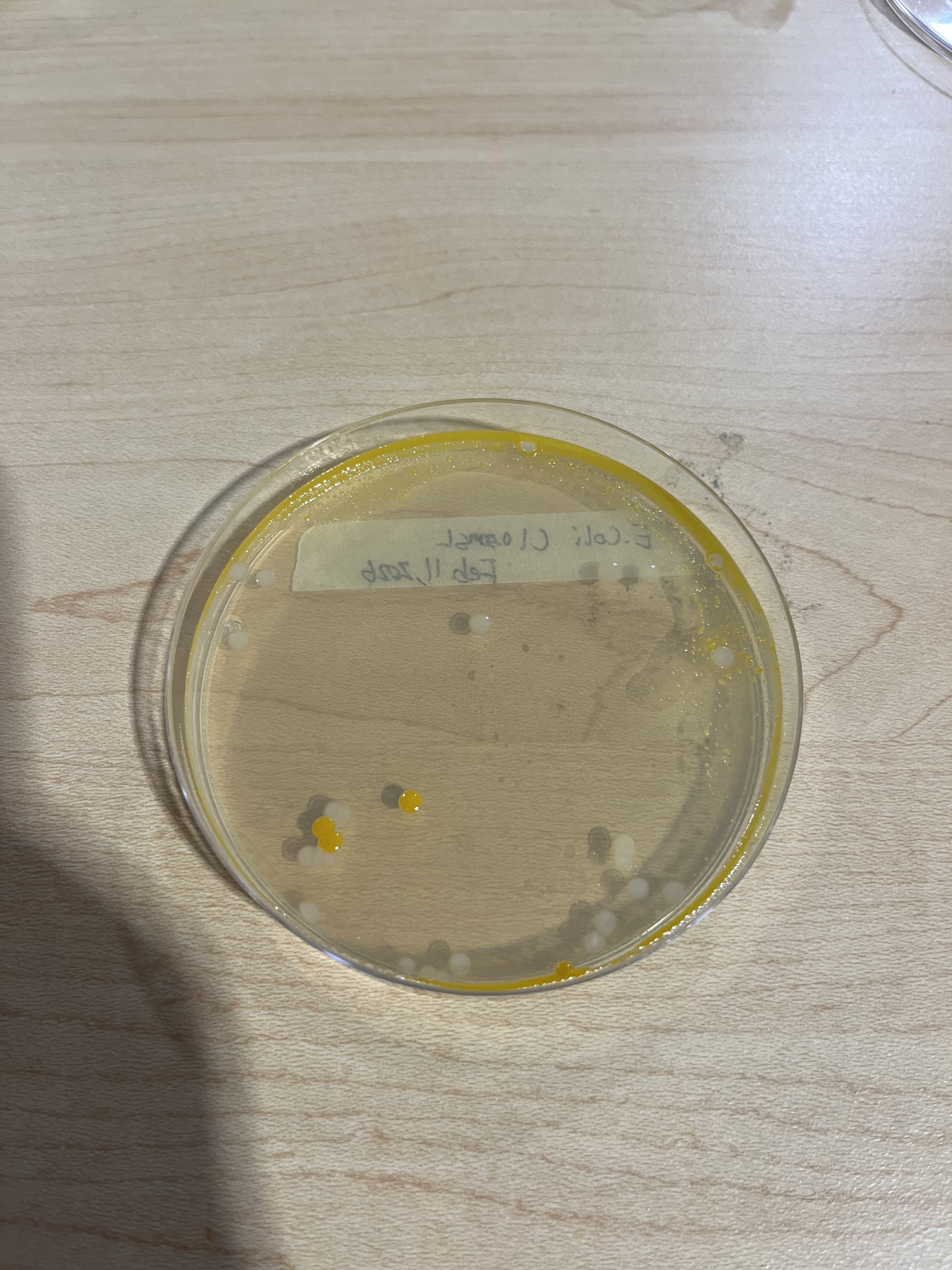
Figure 6: Feb 11th Control Plate, showing the initial start of contamination
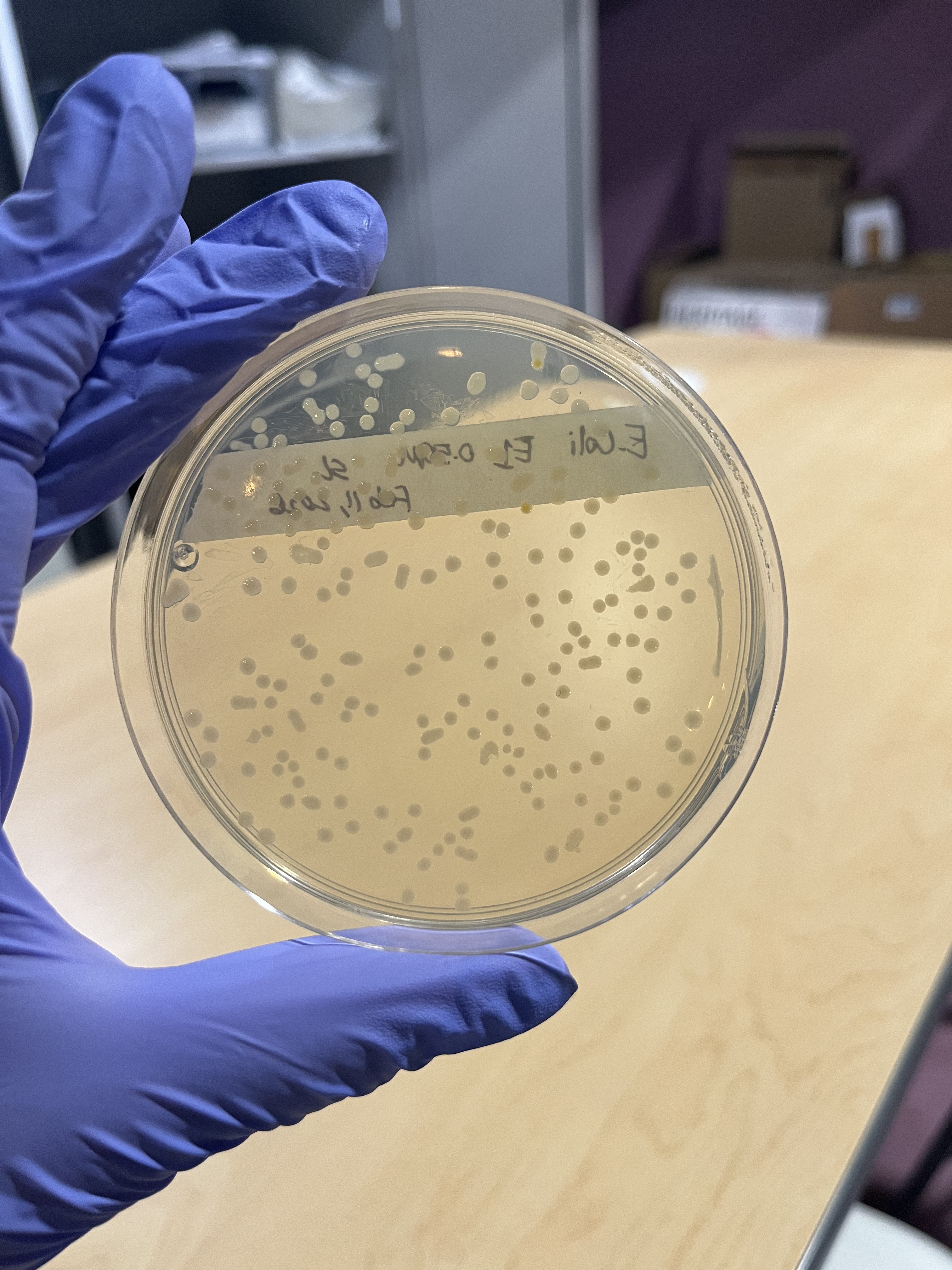
Figure 7: Feb 11th 0.50M Plate after two days of incubation, showing white colonies as E.coli should grow

Figure 8: Feb 24th 0.15M Plate, showing overgrowth of colonies and spread of contamination, which was spread through the rest of experimental data
Discussion
There were many sources of error encountered in this study. During the initial weeks, there was an error in the dilution of E.coli suspensions resulting in overgrowth of bacteria after plating. Furthermore, a contamination issue arose that caused the first serial transfer attempt to result in completely yellow colonies on all plates. This had no reasonable explanation, since all of the experimental and control plates resulted in white mixed with yellow colonies, proving that it was not an error with the magnesium solution. It was assumed that the change might be a shift in the phenotypic expression of the E. coli bacteria in general, however the control plate from the previous week grew white colonies once more. Hence, there was no reasonable explanation reached for the color change of colonies in this experiment.
Another interesting error to mention was the fine balance between the volume of magnesium sulphate stock solution with the volume of agar, as the plates were not able to solidify when solution and agar was at a 10:15 mL ratio, forming small, translucent crystalized suspensions when mixed. Looking at the results, there was an error with the plating and solidification of agar plates in general. In some trials, the agar was not hot enough to form a homogeneous solution with the magnesium, and the resulting plate poured had an uneven surface that may have affected the formation of E.coli colonies. This was especially evident in the plates from the transfer of buffer concentrations to 0.50M during the last week of experiments, explaining the discrepancy in the CFU counts between different trials.
The reason buffer weeks were introduced was to weigh its results against directly transferred bacteria and see if an extra week of serial transfer will help adjust bacterial colonies to the influence of magnesium and increase its survival rates in a higher concentration. Looking at the results, the introduction of a buffer week did significantly increase bacterial survival rates, with the colonies from transferred weeks being more conjugated and smaller in size compared to colonies that were directly transferred to a higher concentration.
Further research could expand upon this study by introducing longer exposure periods of bacteria to magnesium to determine whether sustained or repeated transfer at intermediate magnesium levels could enhance tolerance over time. Long-term serial transfer across multiple generations would also allow for the investigation of potential adaptivity rather than short-term survival alone. Additional studies could also include molecular analysis of bacterial gene responses, providing insight into underlying cellular mechanisms observed by growth patterns.
A closely related study to extend this research would be to increase the magnesium concentrations to which buffered generations of E. coli are exposed. While this study goes up to 0.50M of magnesium sulphate, data at higher concentrations would better define a threshold for bacterial tolerance on solid media. Comparing abrupt exposures to gradual, buffered generations would also share insight into whether stepwise changes can influence survival differently than direct high-level stress exposures.
Finally, this study can be extended into other bacterial species such as potential extremophilic organisms, and even higher-level organisms such as worms and earth-dwelling species to broaden our understanding of organism tolerance in extreme chemical environments.
References
Crisler, J.D., et al. “Bacterial Growth at the High Concentrations of Magnesium Sulfate Found in Martian Soils.” Astrobiology, vol. 12, no. 2, Feb. 2012, pp. 98–106, https://doi.org/10.1089/ast.2011.0720. Accessed 11 Oct. 2020.
Lusk, J E, et al. “Magnesium and the Growth of Escherichia Coli.” Journal of Biological Chemistry, vol. 243, no. 10, May 1968, pp. 2618–2624, https://doi.org/10.1016/s0021-9258(18)93417-4. Accessed 28 Mar. 2023.
Nepal, Sudip, and Pradeep Kumar. “Growth, Cell Division, and Gene Expression of Escherichia Coli at Elevated Concentrations of Magnesium Sulfate: Implications for Habitability of Europa and Mars.” Microorganisms, vol. 8, no. 5, 27 Apr. 2020, p. 637, https://doi.org/10.3390/microorganisms8050637.
Scheller, Eva L., et al. “Formation of Magnesium Carbonates on Earth and Implications for Mars.” Journal of Geophysical Research: Planets, vol. 126, no. 7, July 2021, https://doi.org/10.1029/2021je006828.
Walker, Graeme M. “The Roles of Magnesium in Biotechnology.” Critical Reviews in Biotechnology, vol. 14, no. 4, Jan. 1994, pp. 311–354, https://doi.org/10.3109/07388559409063643.
Wendel, Brian M, et al. “A Central Role for Magnesium Homeostasis during Adaptation to Osmotic Stress.” MBio, vol. 13, no. 1, 15 Feb. 2022, https://doi.org/10.1128/mbio.00092-22.