Soren Bjornson – Life Science, Year 2
Abstract
Growing fresh food is crucial for long space missions but carries significant food safety risks due to the rogue bacteria often present in the closed environment of spacecrafts. Understanding how microgravity affects the interaction between plants and these bacteria, particularly human pathogens, is essential for astronaut safety, as current knowledge is limited. Preliminary research suggests microgravity might weaken plant defences, while increasing the effectiveness of pathogens like E. coli. This study aimed to investigate whether modelled microgravity analogs (MMA) affect the ability of Escherichia coli (E. coli) K-12 to enter the stomata of Lactuca Sativa L. Black Seeded Simpson lettuce. We hypothesized that lettuce plants exposed to MMA would exhibit larger stomatal apertures upon E. coli inoculation compared to plants under normal gravity, allowing for easier bacterial entry. Two inoculated plants were subjected to MMA using a 2D clinostat rotating at 3 RPM for 1 and 3 hours, while three inoculated controls were left vertically under normal gravity for 0, 1, and 3 hours. Epidermal peels were obtained using a tape-peel technique, and stomatal apertures were measured via light microscopy. Our findings show that lettuce plants inoculated with E. coli and exposed to MMA generally exhibited larger average stomatal apertures compared to those maintained under normal gravity conditions. These results show that MMA conditions impair stomatal closure mechanisms in lettuce despite the presence of E. coli, indicating a potential food safety concern for astrobotany. It is suggested that further research into plant-pathogen interactions in microgravity environments be conducted.
Introduction
Modern space travel presents a unique challenge for human sustenance. Long-duration missions require reliable access to nutritious food. While pre-packaged meals can sustain astronauts for a limited time, a study by Cooper et al. (2017) suggests their nutritional value degrades over time, with vitamins such as B1 and C degrading to unacceptable levels after one and three years. The study goes on to state that potassium, calcium, vitamin D, and vitamin K concentrations seem not to meet the recommended daily intake requirements. In our current situation, spaceflight missions over one year are not possible due to these nutritional challenges. Growing leafy greens in space offers a promising solution to this hurdle, with microgreens – the young, fast-growing seedlings of edible vegetables– emerging as a potential source of vitamins and concentrated nutrients. Totsline et al. (2023) suggest that supplementing astronaut’s diets with fresh produce can also help reduce health risks associated with spaceflight, including reductions in bone density, reduced eye health, poor access to vitamin D, reduced uptake of calcium, oxidative stress via cosmic radiation, increased anxiety, and depression, and immunosuppression. However, the closed environment of a spacecraft raises some concerns about food safety. Multiple species of human pathogens have been discovered on surfaces and in the air throughout the International Space Station (ISS), including foodborne pathogens such as Escherichia coli (E. coli), Salmonella enterica, Staphylococcus aureus, and Shigella sonne (Urbaniak et al., 2018). Although no human pathogens have been found in space-grown produce thus far, fungi and other bacteria have, especially in root tissue (Khodadad et al., 2020). Little work has been done on how human pathogens interact with plants under both spaceflight microgravity and modelled microgravity analogs (MMA).
A review by Totsline et al. (2023) highlights the apparent vulnerability of plants to human bacterial pathogens in microgravity. Under both spaceflight microgravity and MMA, plants express their pathogen-defence genes differently, while human bacterial pathogens show increased virulence, antibiotic resistance, stress tolerance, and a reduction in the dose required to be lethal. This suggests an environment where bacteria are more aggressive and plants are less able to defend themselves. Totsline et al. (2023) then go on to explain that, although a universal bacterial response to microgravity is unknown due to the few studies and varying data points, there are still some potential reasons. It is unlikely that gravity alters the intracellular processes of bacteria directly; rather, the altering of their access to oxygen, phosphate, and iron may instead be driving their increased virulence (Pollard, 1965; Totsline et al., 2023). The iron scavenging of human pathogens appears to be enhanced under microgravity, while the lower oxygen and phosphate levels seem to mimic the host environment, triggering the expression of certain virulence genes (Bains et al., 2012; Marteyn et al., 2010; Totsline et al., 2023).
Plants have a relatively rudimentary sense of gravity, employing specialized cells known as statocysts to determine gravity. Each cell contains a number of starch amyloplasts, of which the cell can detect their sedimentation (Kiss, 2010). Under true microgravity, the starch amyloplasts float within the cell, making the plant unable to determine which way is up or down (Totsline et al., 2023). MMA are normally achieved through the use of a 2D or 3D clinostat. These devices slowly rotate at 1-4 rotations per minute (RPM) in either one or both axes, respectively (Thombre et al., 2022). This causes the starch amyloplasts to be in constant free fall, effectively simulating microgravity in plants (Thombre et al., 2022).
Totsline et al. (2023) further note that the specific reaction of plants to true microgravity and MMA varies greatly between different species and cultivars. Many possible differentially expressed genes can occur in response to both types of microgravity, including those associated with reactive oxygen species, cytoskeletal modification, cell-wall development, carbohydrate metabolism, DNA repair, and differential expression of pathogen-defence genes, among others. The review continues, stating how almost half of all upregulated differentially expressed genes were related to wounding and pathogen defence. It has been shown that plants under MMA seem to be unable to close their stomata, keeping them open under both 24-hour light and dark conditions (Treesubsuntorn et al., 2020). As Salmonella and E. coli are unable to break down the cellulose of plant cells and must rely on the pre-existing openings in plant tissue for their entry, the inability to close their stomata could leave plants more susceptible to pathogen colonization.
A study by Totsline et al. (2024) demonstrated how MMA can help facilitate the colonization of Lactuca sativa L. (lettuce) by Salmonella enterica. Their experiment, done under MMA in a clinostat rotating at one to four RPM, revealed wider, more open stomata and deeper bacterial ingression in plants exposed to simulated microgravity when inoculated with Salmonella enterica. This suggests that microgravity weakens a plant’s natural defences, allowing bacterial pathogens to colonize a plant’s intercellular space easily. Other studies were able to show similar trends with non-human and fungal pathogens. Ryba-White et al. (2001), for example, observed an increase in the susceptibility of Glycine max to Phytophthora sojae (the soybean root rot pathogen) under true, in-flight microgravity conditions. Both a greater number of symptoms and more extensive colonization were shown when compared to ground samples. This susceptibility was possibly due to the increased ethylene levels observed in spaceflight samples.
Understanding how true microgravity influences human pathogens in space-grown produce, especially microgreens, through the use of MMA, is essential for developing strategies to ensure that space-grown food is fit for human consumption. By identifying which and how human pathogens exploit the vulnerabilities of plants under microgravity, we can develop mitigation strategies to keep astronauts safe. A potential infection on the ISS or any other spaceship could be devastating, harming astronauts, causing mission delays, and wasting money. This research holds significant implications for ensuring the success of future long-term space missions and the well-being of astronauts throughout them.
While research confirms a potential increase in bacterial colonization under microgravity, the specifics of how human pathogens interact with plants under microgravity are still unknown. Existing studies primarily focus on plant-fungal interactions. My project aims to address these knowledge gaps by investigating how simulated microgravity affects E. coli’s ability to enter Lactuca Sativa L. Black Seeded Simpson lettuce through the stomata. A 2D clinostat will be used to simulate microgravity and compare the colonization rates of E. coli in plants via the opening size of stomata under simulated microgravity versus normal gravity conditions.
Materials and Methods
Plant Material and Growth Conditions:
Lactuca sativa L. var. Black Seeded Simpson seeds (Hunters Gardening Center) were sown in Organic Seed Starting Mix (PRO-MIX Gardening) within disposable plastic seedling trays (Dollarama). Seedlings were cultivated for 17 days in a controlled environment at 20°C under an 84 LED full-spectrum plant grow light (Aigumi). Water was provided every 3 days. Six seedlings that experienced the most growth out of the group were selected for the experiment. The selected seedlings were carefully removed from the soil and rinsed with filtered water to remove any remaining dirt before being surface sterilized by immersion in 70% ethanol for 5 minutes. Sterilized seedlings were placed individually in 8 cm diameter Petri dishes using sterile disposable forceps and secured upright using micropore tape.
Bacterial Culture Preparation:
A single colony-forming unit of E. coli K-12 was moved using a sterile loop into 25 ml of Luria broth to form a liquid culture.The culture was then incubated overnight on an orbital shaker at 100 rpm, 37°c.After incubation, a bacterial pellet was obtained by centrifugation at 4000 rpm for 5 minutes.The supernatant was then asphyxiated, and the bacteria pellet resuspended in a saline solution via vortexing.This washing procedure was repeated twice more, followed by a final resuspension in saline solution.The bacterial suspension was used immediately after preparation.
Inoculation Procedure:
Six wide-tip brushes (Dollarama) were sterilized by immersion in 70% ethanol for one minute, followed by three 10-second rinses in filtered water. The prepared E. coli suspension was vortexed, and a brush was submerged in the suspension for 5 seconds. The abaxial side of each leaf on a seedling was gently brushed with the bacterial suspension, each plant reliably producing three to four leaves. This process was repeated four more times, using fresh brushes and seedlings, as well as vortexing the bacterial suspension each time, resulting in five inoculated plants. A single control plant was treated with filtered water using the same brushing technique. Petri dishes were sealed with micropore tape after inoculation.
Clinostat Experiment:
A 2D clinostat was constructed using two 12v 10 RPM 16ga gear motors operated at 3.5v to achieve a rotation speed of 3 RPM. This specific RPM was chosen to match that of other similar experiments. The motors were adhered with double-sided tape to a wooden mount for stability. Two inoculated seedlings were vertically mounted onto the clinostat using double-sided tape. Samples were collected after one and three hours of rotation. Three inoculated control seedlings (0 RPM) were maintained in a vertical position for zero, one, and three hours, being sampled after their designated time frame. A single uninoculated control seedling (0 RPM) was sampled at zero hours.

Figure 1: A 2D clinostat, as seen during the first trial.
Stomatal Peel Preparation and Microscopy:
Following exposure for the designated time periods, one leaf per seedling was excised using forceps and scalpels sterilized for 5 minutes in 70% ethanol. To assess stomatal apertures, stomata peels were prepared using a modified tape-peel technique (Lawrence et al., 2018). The adaxial leaf surface was first adhered to masking tape (Scotch) for support. Clear cellophane tape (Scotch) was then applied to the abaxial surface and left to sit for five seconds before being carefully peeled off to remove the epidermal layer. The tape, bearing the epidermal layer, was then mounted onto a microscope slide. Stomatal apertures were measured at 500x magnification using a confocal light microscope and a calibrated reticle. Five randomly selected stomata were measured per sample.
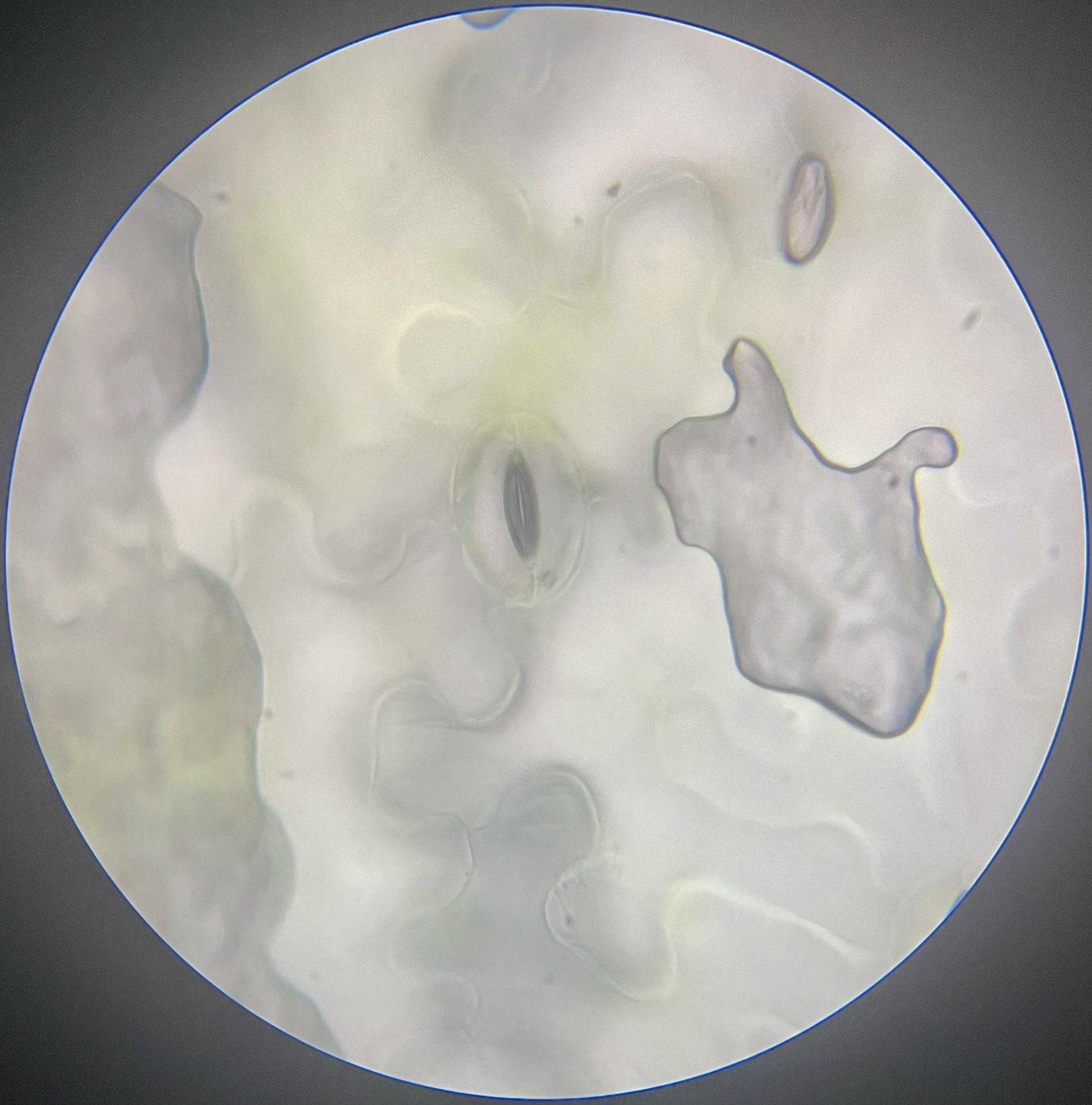
Figure 2: Lettuce stomata inoculated with E. coli for 0 hours viewed under 500x magnification.
Results
To test how E. coli K-12’s ability to colonize lettuce is affected by the presence of MMA, the abaxial sides of Black Seeded Simpson lettuce leaves were inoculated with E. coli suspended in a saline solution. Three of these plants were left in a vertical position for zero, one, and three hours, while two were placed in a clinostat rotating at three RPM for one and three hours. After their designated time, five stomata from each plant were measured, then averaged. This process was then repeated for a second trial. Our results show that E. coli-inoculated lettuce plants under MMA tend to experience larger stomata openings when compared to those not under MMA (Fig. 3). The stomata apertures of one-hour samples under MMA increased in size by an average of 9.8µm compared to those under normal gravity, with the three-hour samples experiencing a difference of 7.7µm (Fig. 3). A notable exception to this lies in the three-hour samples of trial one, where these findings seem to be reversed.

Figure 3: Compares the mean stomatal aperture size (µm) of lettuce leaves across Trials 1 and 2. Compares a mock-inoculated control (vertical, 0 RPM) with inoculated leaves maintained either vertically (0 RPM) or rotated (3 RPM), measured at 1- and 3-hours post-inoculation.
Discussion
The results of this research suggest that MMA conditions may enhance the ability of E. coli to enter and colonize leafy microgreens. These results raise potential concerns for future operations in astrobotany, which may be more easily compromised by stray bacteria and cause food safety concerns. The observation that stomatal aperture size increases while under MMA, even during a bacterial invasion, aligns with previous studies by Totsline et al. (2024) and Treesubsuntorn et al. (2020). Specifically, Treesubsuntorn et al. (2020) reported that under MMA, stomatal apertures in plants remained open regardless of light or dark conditions. This contrasts with normal gravity conditions, where the stomata of control plants typically close in the dark and open in the light. Furthermore, Totsline et al. (2024) demonstrated that simulated microgravity provided Salmonella with an advantage in colonizing and entering plant tissues compared to control groups.
Several limitations may have influenced the results of this study. Firstly, due to time constraints, only two experimental trials were able to be completed instead of the planned three. Secondly, inconsistencies in the rotation speeds of the clinostat’s gear motors were observed between and during trials. This is likely due to a manufacturing defect, as other potential causes were ruled out. Thirdly, the accuracy of sample measurements may have been affected by human interpretation and potential error. Utilizing an inverted-confocal laser microscope in future studies could provide higher-resolution imaging and more precise pixel-based measurements to address this. Fourthly, the inability to preserve leaf samples before measurement due to safety concerns regarding formaldehyde use could have led to slight variations in stomatal size over the sampling period. Finally, for an unknown reason, the three-hour samples in the first trial wilted and became weak after their designated time. This could explain why those results were outliers. Given these limitations, it is recommended that another research group with more funding, resources, and time replicate this study to validate these findings.
Similar studies could be conducted using a range of different bacteria, viruses, fungi, and plants to confirm that the effects of MMA are similar across species. These interactions could also be examined under true microgravity conditions or using a 3D clinostat. Additionally, future research could focus on gaining a more solid understanding of why MMA affects both plant physiology and pathogen virulence. Further research could also vary the light conditions, experiment with longer durations under MMA, or even use seedlings grown entirely under MMA conditions.
This study’s findings suggest that MMA conditions can increase the susceptibility of plants to bacterial pathogens, specifically by inhibiting the closure of stomata. These findings align with existing papers demonstrating that MMA can hinder stomatal closure despite lighting conditions and facilitate the entry of bacterial pathogens like Salmonella (Totsline et al., 2024; Treesubsuntorn et al., 2020). The implications of this research are relevant to the field of astrobotany, raising concerns about the potential food safety issues in future long-duration space missions, where the cultivation of fresh produce might be required for astronaut health and nutrition.
References
Bains, M., Fernández, L., & Hancock, R. E. W. (2012). Phosphate Starvation Promotes Swarming Motility and Cytotoxicity of Pseudomonas aeruginosa. Applied and Environmental Microbiology, 78(18), 6762–6768. https://doi.org/10.1128/aem.01015-12
Cooper, M., Perchonok, M., & Douglas, G. L. (2017). Initial assessment of the nutritional quality of the space food system over three years of ambient storage. Npj Microgravity, 3(1). https://doi.org/10.1038/s41526-017-0022-z
Khodadad, C. L. M., Hummerick, M. E., Spencer, L. E., Dixit, A. R., Richards, J. T., Romeyn, M. W., Smith, T. M., Wheeler, R. M., & Massa, G. D. (2020). Microbiological and nutritional analysis of lettuce crops grown on the International Space Station. Frontiers in Plant Science, 11. https://doi.org/10.3389/fpls.2020.00199
Kiss, J. Z. (2010). Mechanisms of the early phases of plant gravitropism. Critical Reviews in Plant Sciences, 19(6), 551–573. https://doi.org/10.1080/07352680091139295
Lawrence, S., II, Pang, Q., Kong, W., & Chen, S. (2018). Stomata Tape-Peel: an improved method for guard cell sample preparation. Journal of Visualized Experiments, 137. https://doi.org/10.3791/57422
Marteyn, B., Scorza, F. B., Sansonetti, P. J., & Tang, C. (2010). Breathing life into pathogens: the influence of oxygen on bacterial virulence and host responses in the gastrointestinal tract. Cellular Microbiology, 13(2), 171–176. https://doi.org/10.1111/j.1462-5822.2010.01549.x
Pollard, E. C. (1965). Theoretical studies on living systems in the absence of mechanical stress. Journal of Theoretical Biology, 8(1), 113–123. https://doi.org/10.1016/0022-5193(65)90097-4
Ryba-White, M., Nedukha, O., Hilaire, E., Guikema, J. A., Kordyum, E., & Leach, J. E. (2001). Growth in microgravity increases susceptibility of soybean to a fungal pathogen. Plant and Cell Physiology, 42(6), 657–664. https://doi.org/10.1093/pcp/pce082
Thombre, R. S., Kaur, K., Jagtap, S. S., Dixit, J., & Vaishampayan, P. V. (2022). Microbial life in space. In R. Thombre & P. Vaishampayan (Eds.), New Frontiers in Astrobiology. Elsevier. https://doi.org/10.1016/B978-0-12-824162-2.00013-0
Totsline, N., Kniel, K. E., & Bais, H. P. (2023). Microgravity and evasion of plant innate immunity by human bacterial pathogens. Npj Microgravity, 9(1). https://doi.org/10.1038/s41526-023-00323-x
Totsline, N., Kniel, K. E., Sabagyanam, C., & Bais, H. P. (2024). Simulated microgravity facilitates stomatal ingression by Salmonella in lettuce and suppresses a biocontrol agent. Scientific Reports, 14(1). https://doi.org/10.1038/s41598-024-51573-y
Treesubsuntorn, C., Lakaew, K., Autarmat, S., & Thiravetyan, P. (2020). Enhancing benzene removal by Chlorophytum comosum under simulation microgravity system: Effect of light-dark conditions and indole-3-acetic acid. Acta Astronautica, 175, 396–404. https://doi.org/10.1016/j.actaastro.2020.05.061
Urbaniak, C., Sielaff, A. C., Frey, K. G., Allen, J. E., Singh, N., Jaing, C., Wheeler, K., & Venkateswaran, K. (2018). Detection of antimicrobial resistance genes associated with the International Space Station environmental surfaces. Scientific Reports, 8(1). https://doi.org/10.1038/s41598-017-18506-4